- Les données issues des mouillages
Les séries temporelles haute fréquence des variables mesurées nous renseignent sur la variabilité temporelle du milieu et nous permettent d'extraire des tendances long terme. Les Figures 8 à 12 montrent des exemples très révélateurs de ce qu'il est possible de mesurer avec ces plateformes fixes. La figure 8 montre par exemple comment il est possible grâce aux mouillages LION et DYFAMED, d'étudier la variabilité interannuelle du phénomène de convection profonde. Les années 2008 et 2014-2018 sont marquées par une faible convection profonde tandis qu'elle a atteint le fond les autres hivers. De 2009 à 2013, la convection profonde montre les mêmes caractéristiques en intensité à quelques décalages temporels et en particulier un second épisode convectif après une première phase de mélange violent qui atteint le fond (Houpert et al 2016). L'hiver 2018 (février-mars) fut également caractérisé par un fort mélange qui a surtout impacté la LIW (Margirier et al., 2020). On peut aussi observer des variabilités différentes suivant les deux sites . Par exemple, de part et d'autre du bassin on observe une nette accentuation du contenu thermique et halin de la LIW (300-800m) de 2014 à 2018 alors qu'on observe un impact différent sur le contenu en O2 (Fig. 9). En effet, alors que les concentrations en O2 au site LION sont relativement bien ventilées chaque hiver de 2014 à 2018 (la MLD atteint souvent 300m sur ce site), elles ne le sont pas au site DYFAMED, situé dans une zone moins soumise au forçage des vents froids et secs en hiver et donc à un mélange profond pouvant atteindre la LIW. De ce fait, on observe très nettement une diminution des concentrations en O2 caractéristique vraisemblablement d'une activité bactérienne (respiration).
Le maillage effectué dans le bassin nous renseigne essentiellement sur des contrastes Est-Ouest entre la mer Ligure et le Golfe du Lion et les signaux enregistrés sont largement dominés par le cycle saisonnier, les variations de méso-échelle et les ondes internes. Dans ces enregistrements, une variabilité de quelques jours à quelques semaines domine le signal. Un problème majeur avec les données de mouillage est qu'on peut difficilement décorréler le signal temporel du signal spatial. Dans quelle mesure une variation enregistrée au point fixe peut-elle être représentative d'une échelle spatiale? Les mesures de courants nous permettent d'avoir une idée de la présence de structures dynamiques comme des tourbillons. Cependant, il est plus difficile d'extrapoler sur l'échelle de l'ensemble du bassin sans données complémentaires. En se focalisant sur certaines échelles et en faisant une analyse soignée du signal, il est possible de faire émerger certains signaux pour les variables mesurées.

Figure 8 : Évolution de la température aux sites DYFAMED (haut) et LION (bas) de 2007 à 2018. La ligne noire représente la profondeur de la couche de mélange (ou MLD) qui indique l'intensité de la convection profonde en hiver (Margirier et al., 2020).
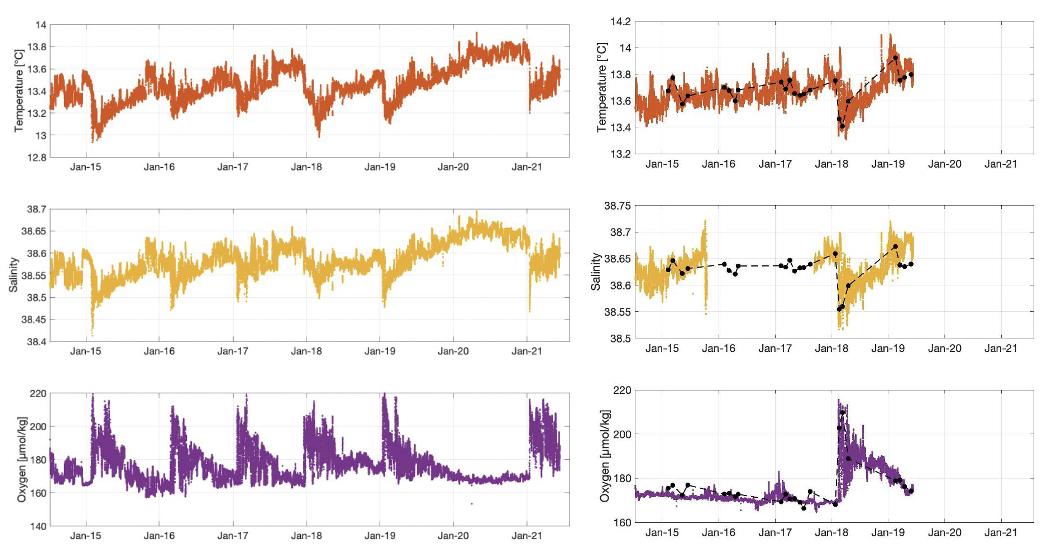
Figure 9 : Exemple de données T, S et O2 enregistrées sur le mouillage LION (droite) et DYFAMED avec les données issues des campagnes mensuelles (gauche) de juin 2014 à juin 2021 à 300m (niveau de la LIW). Les mesures du mouillage à DYFAMED 2019-2021 ne sont pas montrées ici en raison de la perte du mouillage de décembre 2019 à juin 2021
Les figures 10 et 11 montrent des résultats issus des analyses des concentrations et flux particulaires à LION et à DYFAMED que l'on peut corréler avec les séries temporelles de courants horizontaux et verticaux et de température ainsi que la variabilité de la profondeur de la couche de mélange. Au site DYFAMED, on observe également que les flux de masse diminuent depuis les années 2000 alors que le contenu en carbone organique (%OC) augmenterait à 200m et diminuerait à 1000m (Fig. 11). Cette inversion dans la tendance s'expliquerait par un export de carbone sous la couche euphotique plus riche en carbone labile ces dernières années alors que la boucle microbienne (et donc dégradation de carbone) serait plus efficace et/ou active dans la zone obscure (entre 200 et 1000m).
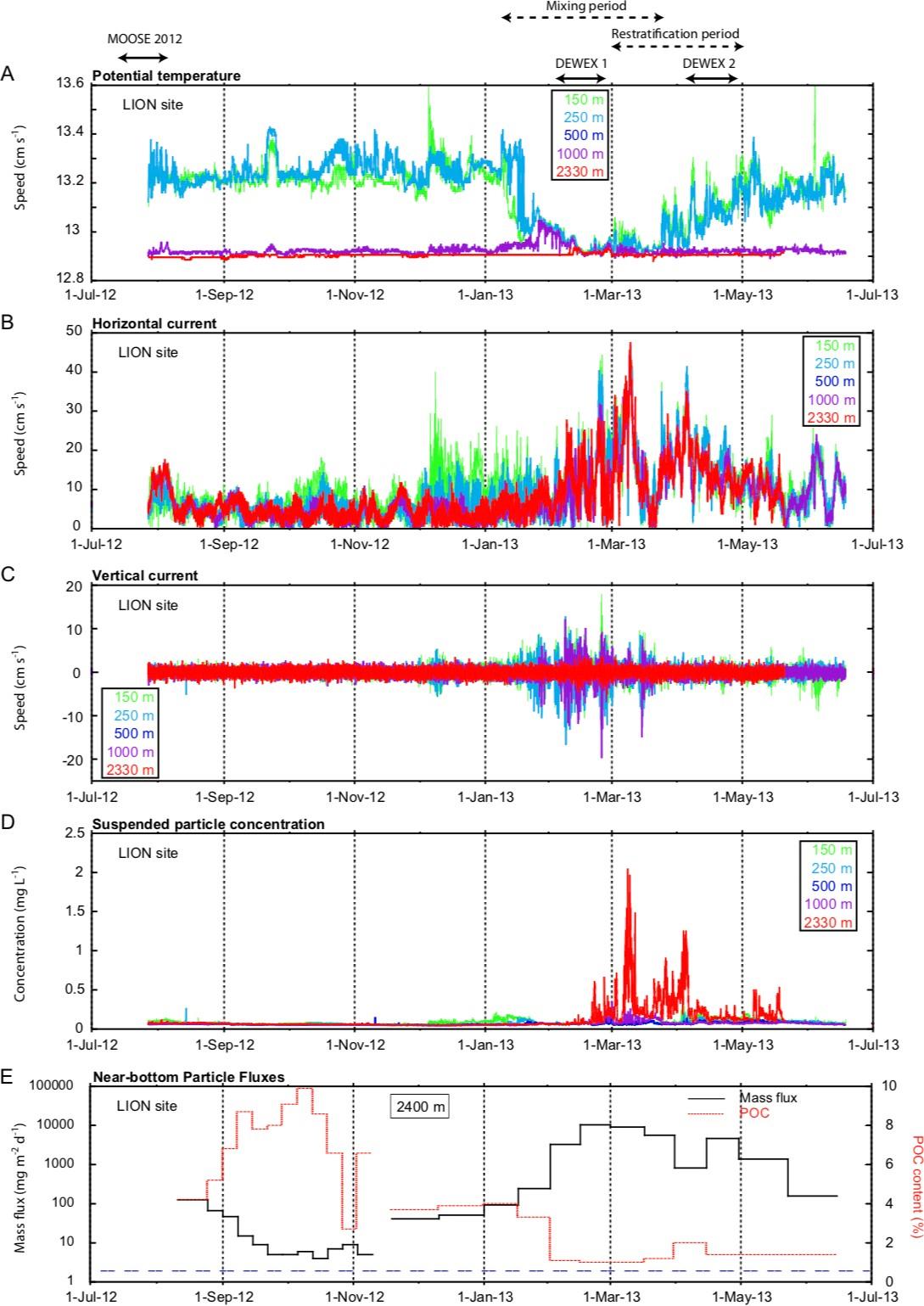
Figure 10 : Évolution de la température potentielle, des vitesses horizontales et verticales des courants et de la concentration en particules (dérivée de l'intensité acoustique rétrodiffusée) mesurées par les dans la colonne d'eau par les différents courantomètres sur la ligne LION. La figure du bas montre l'évolution à proximité du fond du flux de masse et la teneur en carbone organique mesurée par le piège à particules à la ligne LIONCEAU. Ces observations révèlent l'effet de la convection profonde sur l'augmentation des flux particulaires liée à la resuspension de sédiments profonds pauvres en carbone organique particulaire (Durrieu de Madron et al., 2017).
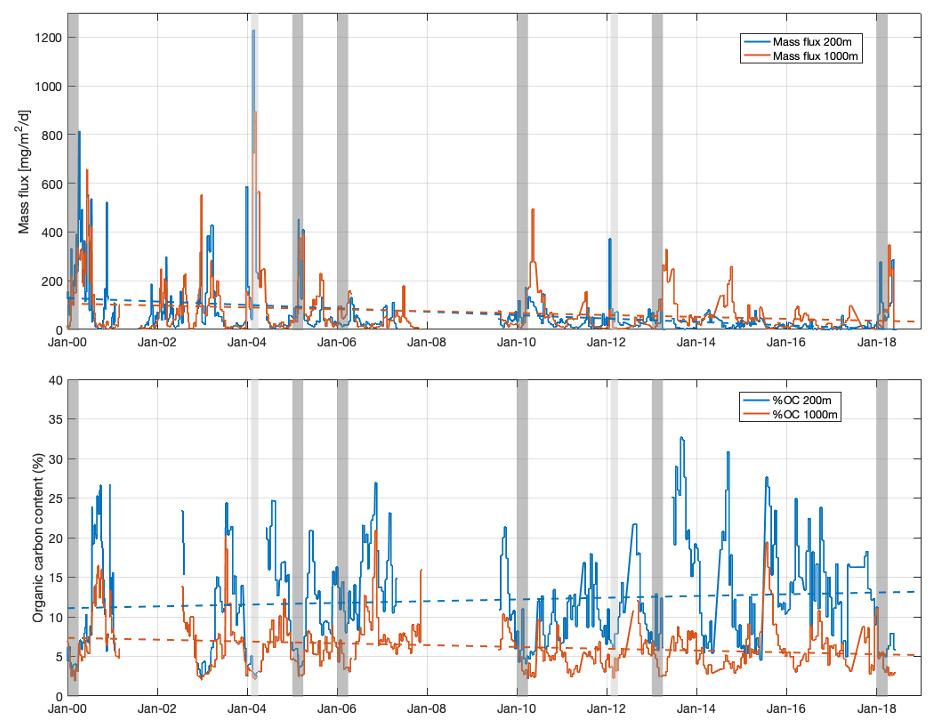
Figure 11 : Série temporelle des flux de masse et de carbone organique au site DYFAMED à 200 et 1000m (les barres gris foncé et gris clair indiquent respectivement les années de convection profonde avec une MLD inférieur à 1000 m et les années moins convectives avec une MLD autour de 200 m). La variabilité des flux à 200m est principalement impactée par l'intensité du mélange qui a lieu en hiver, comme par exemple en hiver 2012, 2013 et 2018 marqué par une forte convection en mer Ligure. Les flux à 1000m dépendent également de l'intensité de cette convection mais également des processus d'agrégation qui ont lieu lors de l'export des particules dans la zone mésopélagique.
La figure 12 montre l'évolution de la température des eaux levantines intermédiaires dans le courant Nord sur une trentaine d'années au niveau du canyon de Lacaze-Duthiers dans le Golfe du Lion. La forte variabilité saisonnière et les chutes de température en période hivernale résultent du débordement et de la plongée le long de la pente continentale des eaux froides et dense formées sur le plateau. Une tendance visible au réchauffement sur le long terme au niveau des eaux intermédiaires est visible depuis 2014. Les derniers évènements de cascades profondes, ayant un effet de refroidissement sur les caractéristiques des eaux de fond de Méditerranée occidentale datent des hivers 2012 et 2013.

Figure 12 : Séries temporelles de température à mi-hauteur d'eau, à 500 m (A), et près du fond, à 1000 m (B) de profondeur dans le canyon de Lacaze-Duthiers, et près du fond à 2400 m dans la zone de convection profonde au site LION (C).
- Le réseau de stations CTD MOOSE-GE « grande échelle »
Les Figures 13 et 14 montrent des exemples de données récoltées dans le cadre de campagnes MOOSE-GE. La Figure 13 documente bien les différents grands traits de circulation avec le Courant Nord caractérisé par des LIW bien marquées qui s'écoulent le long du talus, le passage des eaux profondes nouvelles et de WIW (minimum de température) au nord de Minorque et le Gyre Nord dont le centre est caractérisé par des LIW un peu moins marquées car elles ont été érodées par les phénomènes de formation d'eau en hiver.
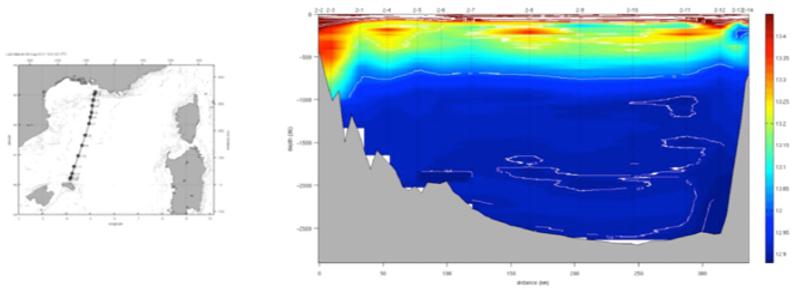
Figure 13 : Section de température potentielle de la campagne MOOSE-GE 2012 en fonction de la distance à la station la plus proche de Marseille.
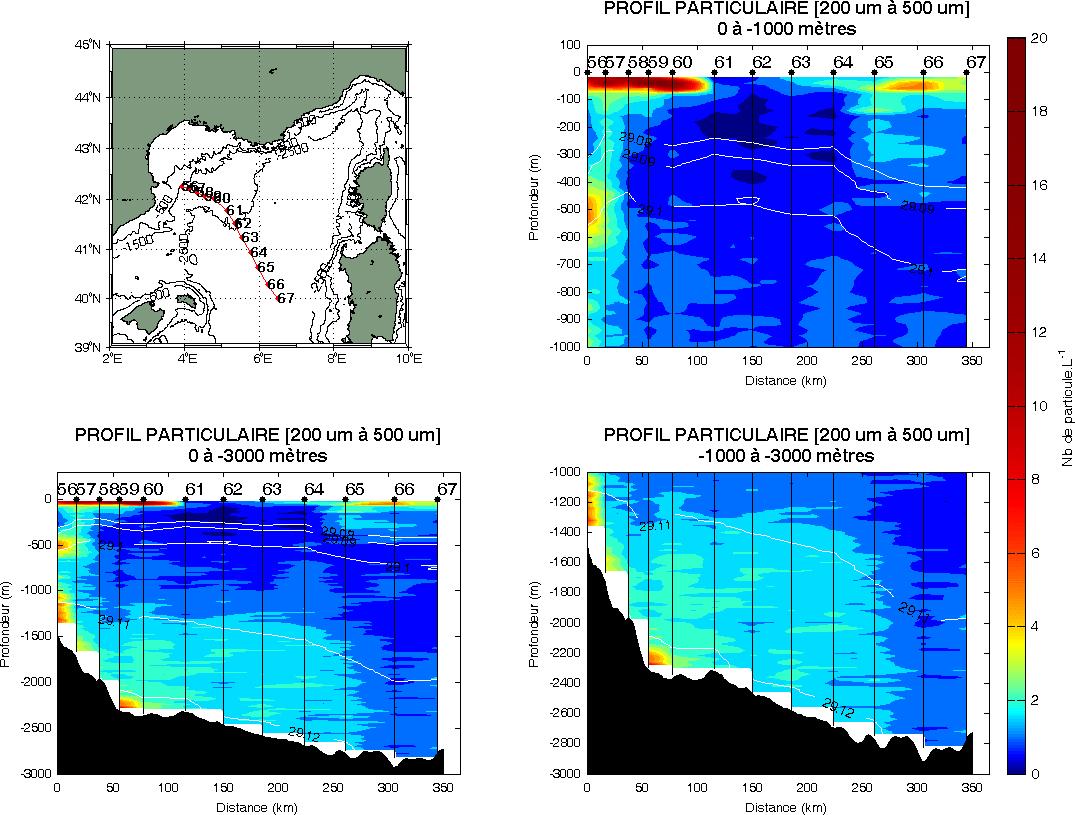
Figure 14 : Exemple de radiale UVP indiquant l'abondance en taille de particules (ici de 200 à 500 µm) sur plus de 200km le long de la radiale Lacaze-Sardaigne en été 2012.
Les mesures d'abondance de particules classées par type de taille (Figure 14) avec l'UVP5 («Underwater Vision Profiler») utilisé pendant les campagnes MOOSE-GE montre une distribution homogène des petites particules (<500 µm) sur toute la colonne d'eau et l'impact sur le long terme (~4 mois) des phénomènes de « cascading» dans le Golfe du Lion. L'action de tourbillons (conséquence d'une restratification hivernale) interagissant avec le fond pourraient expliquer le phénomène de resuspension des petites particules (200m au-dessus du fond) le long de la section Lacaze-Lion.
Ce réseau de stations a aussi permis de cartographier le volume et l'étalement des eaux profondes nouvelles en 2012 (Figure 15). Il est possible d'effectuer ce même genre de bilan pour les autres variables mesurées. On distingue généralement la zone du talus séparé par un front d'une zone au large où les gradients grande échelle sont moins importants.
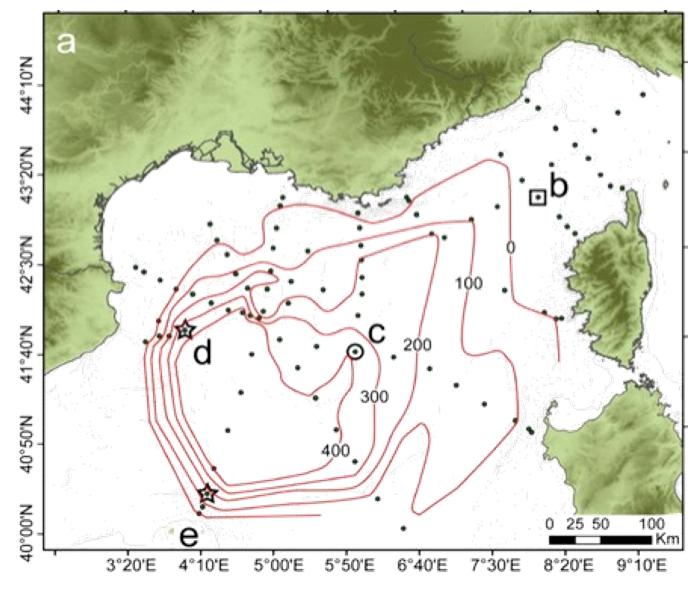
Figure 15 : Cartographie des eaux profondes d'après Durrieu de Madron et al. (2013). Hauteur au-dessus du fond de l'isopycne 29.11 qui retrace bien le volume des eaux profondes nouvelles et leur dispersion, environ 4 mois après les événements de mélange violent hivernal.
Enfin, la série temporelle des données hydrographiques de 2010 à 2014, montrent bien l'évolution de cette nouvelle masse d'eau formée tantôt par la convection au large, tantôt par le phénomène de « cascading » (ou les deux associés), provoquant ainsi un empilement successif des anciennes masses d'eau dense formée les hivers précédents. La nouvelle masse d'eau dense formée pendant les forts processus de convection est identifiable par la forme de crochet dans les diagrammes theta-S-O2 (figure 16).
Le réseau en étoile réparti dans tout le bassin est idéal pour mettre à jours nos climatologies (T, S, courants, mais aussi nutriments et DIC) et il est effectivement utilisé pour initialiser les modèles numériques SYMPHONIE/NEMO et PISCES/ ECO3M (Estournel et al. 2015, Waldman et al. 2015; Ulses et al., 2021). Dans une perspective d'assimilation de données (réanalyse MERCATOR sur la Méditerranée), les campagnes MOOSE-GE fournissent un jeu de données à même de bien contraindre les modèles numériques de prévision. Il a été d'ailleurs montré qu'elles permettaient d'ajuster les états initiaux des modèles en été et ainsi de mieux prédire les épisodes de convection profonde de l'hiver suivant (épisodes mesurés plus intensivement notamment dans le cadre des campagnes DEWEX réalisées sur le Suroît en 2013).
Le jeu de données ainsi acquis est à ce titre unique et devrait constituer une référence en la matière pour les années à venir. Les campagnes MOOSE-GE constituent la majeure partie des mesures disponibles pour la plupart des variables mesurées dans le cadre de MOOSE. Une complémentarité avec la description de la mésoéchelle et le suivi temporel continu est assurée par les autres moyens d'observations de MOOSE (sorties mensuelles, gliders, flotteurs Argo, radars HF, etc.).
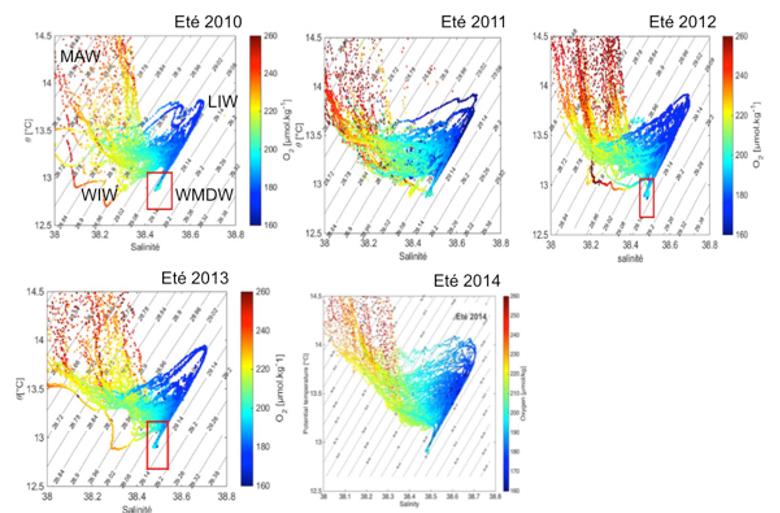
Figure 16 : Évolution des caractéristiques des masses d'eau observées dans le bassin pendant les campagnes MOOSE-GE de 2010 à 2014. Les concentrations du minimum en oxygène (valeur en couleur) révèlent bien le signal de la LIW plus ou moins marqué chaque année. Au fond, le crochet TS-O2 montre la présence de nouvelle masse d'eau dense formée par des eaux plus froide/salée (« cascading ») et/ou plus chaude/salée (convection au large) et plus riche en O2 notamment en 2012/2013.
- Bilan des données permettant de caractériser le système des carbonates océanique.
Les mesures du système des carbonates se font depuis 2015 le long de deux radiales nord-sud (Nice-Calvi et Minorque- Marseille) coupant le front des Baléares et le courant Nord (auparavant les données étaient collectées de part et d'autre du bassin). Ces radiales ont pour avantage de mieux décrire la variabilité interannuelle AT-CT dans différentes masses d'eau, le long d'un continuum côte-large ayant subi un mélange hivernal plus ou moins intense à l'ouest et à l'est de la zone de la Méditerranée Nord-Occidentale (Figure 17). Les mesures AT-CT (et de pHT depuis 2019 ¿ voir partie « DÉMARCHE QUALITÉ ET VALIDATION DES DONNÉES ») lors des campagnes MOOSE-GE permettent d'estimer les autres variables du système des carbonates (comme par exemple la pCO2 de surface ou encore le pH).
L'étude des tendances d'évolution du CO2 dans les masses d'eau permettant de détecter les effets naturels (climat) et humains (réchauffement, acidification) conduisant à un changement sur la chimie des carbonates nécessite un suivi sur un temps suffisamment long pour détecter des signaux significatifs. De ce point de vue, les données acquises au cours des campagnes MOOSE-GE constituent un suivi indispensable, complémentaire des suivies mensuelles au stations fixes du réseau MOOSE, pour extrapoler les tendances observées à des points fixes à l'ensemble du bassin. Mais une première décennie de données acquises ne constitue que les prémices de ces études sur les tendances à long terme. On peut cependant déjà citer des exemples de contributions significatives des données de chimie des carbonates acquises au cours des campagnes MOOSE-GE :
- Les données MOOSE-GE couvrant le bassin liguro-provençale ont permis de mettre en évidence une tendances décennale à l'acidification et un enrichissement du contenu en carbone inorganique de l'ensemble de la colonne d'eau de ce bassin (Coppola et al. 2020).
- L'utilisation du jeu de données de MOOSE-GE pour entraîner un réseau de neurones permettant d'estimer le système des carbonates à partir des données de température, salinité et oxygène dissous. Ainsi à partir des données T, S, O2 des flotteurs Argo et des mouillages, ce réseau CANYON-MED permet d'augmenter la résolution temporelle et spatiale des observations (voir partie «Développement d'un réseau de neurones en Méditerranée entraîné par les données des campagnes MOOSE-GE »)
- Les données MOOSE-GE alimentent de manière significative les bases de données internationales de CO2 océanique dans la colonne d'eau. Les données des campagnes MOOSE-GE sont incluses dans la base de données CARIMED qui vise à qualifier et ajuster les données Méditerranéenne avant de les intégrer dans la base de données GLODAP (www.glodap.info) à l'échelle globale.
- Les données MOOSE-GE contribuent à la réalisation d'un « carbon audit » de la Méditerranée Nord-Occidentale dans le cadre du projet H2020 EUROSEA. Il est à noter que l'importante couverture spatiale que permet le suivi MOOSE-GE a constitué une motivation pour proposer cette zone comme projet pilote de « Carbon Audit » dans le cadre de ce projet.
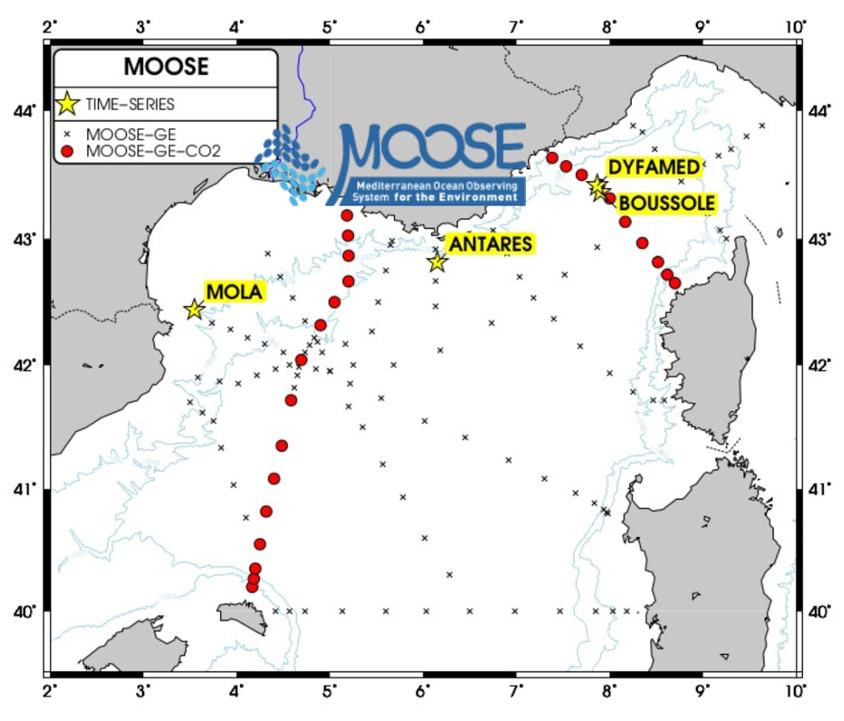
Figure 17: maillage du réseau CO2 dans les campagnes MOOSE-GE. Les deux radiales nord-sud Nice-Calvi et Marseille-Minorque permettent d'échantillonner différentes masses d'eau et notamment l'impact de la convection hivernale de la surface au fond. Ceci permet également de mieux observer l'évolution du CO2-pH dans le système côte-large
- Bilan des données de sels nutritifs
Les données MOOSE-GE ont été utilisées pour actualiser la climatologie des sels nutritifs de la mer Méditerranée occidentale (Figure 178 Belgacem et al., 2021). Des champs moyens annuels de nutriments maillés pour la période 1981 - 2017 et ses sous-périodes 1981 - 2004 et 2005 - 2017 ont été produits sur une grille horizontale ¿° × ¿° (figure ci-dessous). Parmi les données utilisées (MedAtlas, campagnes CNR, MedShip et MOOSE-GE), MOOSE-GE représente 25% des données sur la période 1981 à 2017 sachant que le programme MOOSE a débuté en 2010. La contribution des campagnes représentent ici un atout puisque le maillage des stations est plus dense et la fréquence des campagnes plus élevées offrant ainsi une meilleure représentation spatiale et temporelle depuis 2010. Enfin, les campagnes MOOSE-GE permettent également d'échantillonner davantage les eaux profondes en sels nutritifs puisque les stations sont échantillonnées jusqu'à 10m au-dessus du fond.
Une analyse de sensibilité a été effectuée pour évaluer la comparabilité du produit de données avec les données d'observation. Le nouveau produit fournit une bonne référence pour la région et pour la communauté scientifique pour la compréhension de la variabilité des nutriments inorganiques en Méditerranée occidentale, dans l'espace et dans le temps. Il représente très bien les schémas spatiaux de la distribution des nutriments en raison de sa plus grande couverture de données spatiales et temporelles par rapport aux produits climatologiques existants et contribue à la compréhension de la variabilité spatiale des nutriments en Méditerranée occidentale.
Le nouveau produit capture le fort gradient de nutriments est-ouest et les caractéristiques verticales. Les résultats obtenus ne comprennent pas de champs d'analyse saisonniers ou annuels. Cependant, l'ensemble de données agrégées ici montre des améliorations dans la description de la distribution spatiale des nutriments inorganiques dans la Méditerranée occidentale. Nous reconnaissons que le calcul d'une moyenne climatologique sur une période donnée n'est pas suffisant pour estimer et détecter la tendance induite par le changement de climat. Pour les tendances, la communauté MOOSE s'appuiera en particulier sur les prédictions produites par le réseau de neurones CANYON-MED (Fourrier et al., 2021). Cependant, la comparaison des climatologies basées sur les deux périodes, 1981-2004 (avant le changement climatique mondial) et 2005-2017 (après le changement climatique mondial), a déjà produit des résultats importants. Des changements notables ont été constatés dans la distribution des nutriments après le WMT à différentes profondeurs. Ces résultats confirment la tendance à une augmentation relativement croissante de nutriments inorganiques dans le bassin et établissent un lien possible entre le changement des schémas de la circulation générale, les changements dans la stratification profonde et les tendances au réchauffement induisant un affaiblissement des épisodes de convection en hiver dans le centre du Golfe du Lion.
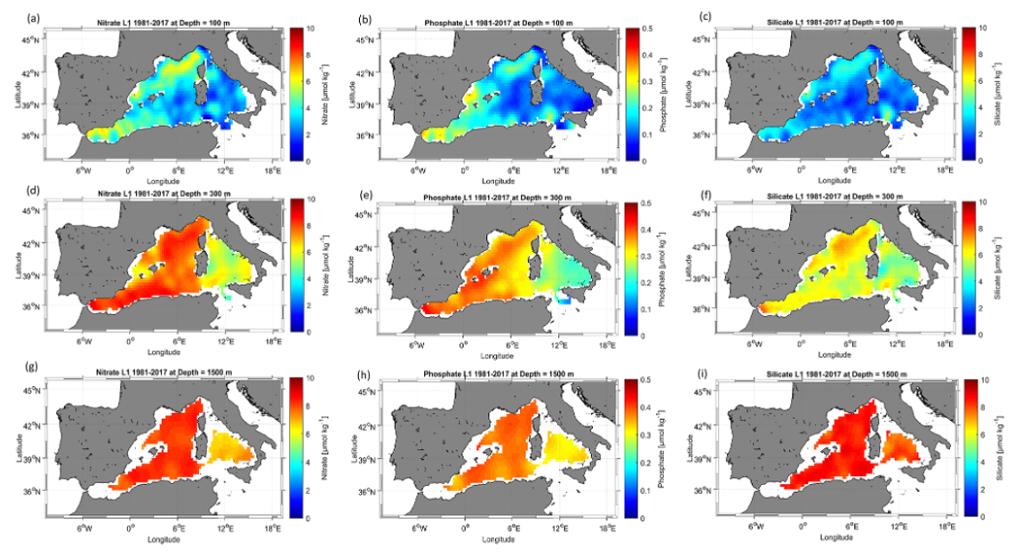
Figure 18 : Cartes de climatologie des nitrates (a- à 100 m, d- à 300 m, g- à 1500 m), des phosphates (b- à 100 m, e- à 300 m, h- à 1500 m) et des silicates (c- à 100 m, f- à 300 m, i- à 1500 m) pour la période de 1981 à 2017.
- Les observations du zooplancton à travers les campagnes MOOSE-GE (imagerie et génomique)
Les données biologiques réalisées lors des campagnes MOOSE-GE depuis 2014, ce sont focalisées principalement sur la diversité zooplanctonique obtenue à partir d'images le long de la colonne d'eau (profileur UVP) et à partir d'échantillons collectés par les traits de filets verticaux (de 15 à 18 stations environ par campagne). Les premiers résultats des échantillons de filets (mesurés par Zooscan) issus des campagnes 2015 et 2016 ont montré que le mésozooplancton épipélagique est concentré dans la couche 0-100m avec une répartition spatiale dépendante des conditions physico- chimiques (Fig. 19). Également la différence dans les données 2015 et 2016 est fortement liée à la période d'échantillonnage avec une campagne en été (juillet 2015) et une en fin de printemps (mai 2016).
En outre, nous avons observé une forte variabilité intra-annuelle dans les communautés qui s'expliquerait principalement par le gradient de latitude. C'est une observation classique en biologie mais il serait intéressant d'étudier cet effet lors d'un passage de front à travers le gradient hydrologique. Il reste encore à coupler ces données aux mesures UVP afin d'étudier quelles espèces seraient mieux adaptées à certaines conditions physico-chimiques et de comprendre pourquoi.
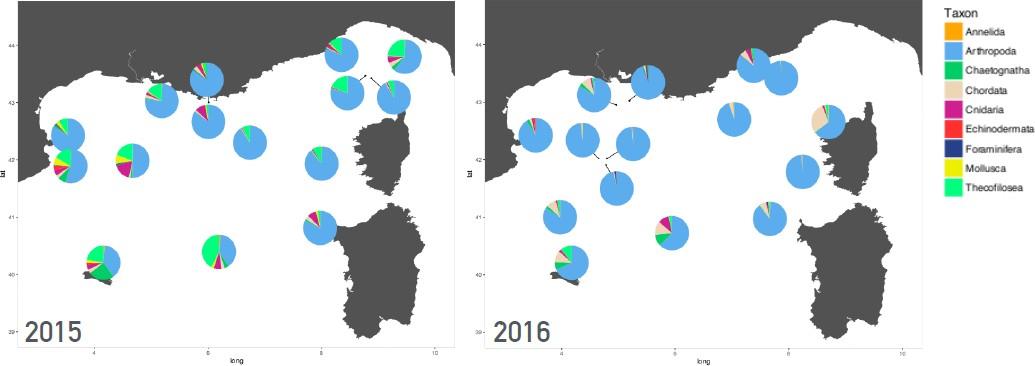
Figure 19 : Graphiques circulaires représentant l'abondance relative du zooplancton dans les échantillons de l'année 2015 et 2016 identifiés avec le ZOOSCAN.
La Méditerranée nord-occidentale est une bio-région à productivité printanière et estivale fortement dépendante du processus de convection hivernale des eaux profondes. En 2017, bien que la convection hivernale profonde ait été d'intensité faible à moyenne et que l'échantillonnage de MOOSE-GE 2017 ait eu lieu en fin d'été (septembre), les schémas de distribution du zooplancton dans la région centrale de Méditerranée nord occidentale et les taux associés estimés ont révélé l'empreinte de ce forçage majeur en fin d'été sur le zooplancton (Fig.20; G. Feliu, 2021). Dans sa thèse, Guillermo Feliu (MIO) présente une analyse détaillée de la structure et du fonctionnement de la communauté zooplanctonique échantillonnée lors de la campagne MOOSE-GE de fin d'été 2017 (Figure 20) et compare ces résultats avec ceux d'autres écosystèmes marins méditerranéens (Campagne PEACETIME en mai 2017), et avec des campagnes antérieures dans la même région liguro-provençale (DEWEX 20013 et MOOSE-GE 2014) (Tableau 5). Pour ce travail, nous avons utilisé différentes méthodes de traitement des échantillons permettant de délivrer des abondances et d'estimer des biomasses : ZOOSCAN et FlowCAM d'une part, et le comptage au microscope d'autre part, et les comparaisons entre ces deux approches ont montré une très bonne similitude dans les gammes de concentrations habituelles de zooplancton en mer Méditerranée (entre 500 et 3000 ind.m-3) dans la couche épipélagiques 0-200m.

Figure 20: Distributions spatiales des variables du zooplancton pendant MOOSE-GE 2017. Abondance (a), biomasse estimée (b), pente NBSS des spectres de taille (c). Indice de diversité de Shannon (d), rapport p1/p2 des rangs-fréquences (e), rapport Adultes/Copépodites (f). (Thèse Feliu , 2021)
Pour MOOSE-GE 2017, trois clusters spatiaux distincts ont été trouvés sur la base des valeurs intégrées 0-200 m des variables environnementales : le premier avec des stations principalement situées dans la Mer Ligure, et les deux autres situées dans les parties nord et sud du Bassin Provençal, mettant en évidence un gradient nord-sud notable. Les abondances et la biomasses (estimée à partir des biovolumes) du zooplancton dans la Mer Ligure et le Bassin Provençal Sud étaient plutôt homogènes spatialement, et les variations les plus importantes ont été trouvées dans le Bassin Provençal central en bordure du plateau du Golfe du Lion. Ces valeurs automnales de 2017, obtenues après un phénomène printanier de convection profonde modeste (Margirier et al., 2020) étaient comparables à des valeurs obtenues dans ces mêmes régions pour la même saison avant les années 2010, également moyennement à peu convectives (sept-oct 2004 : Raybaud et al.,2008, pour la mer Ligure ; été 2008 Nowaczyk et al., 2011, pour le bassin provençal). On a pu observer que les valeurs d'abondance et de biomasse enregistrées pendant MOOSE-GE 2017 au sud du bassin provençal étaient très similaires à celles enregistrées pendant PEACETIME à la mi-mai 2017 environ 1300 ind.m-3 et 3,5 mg DW .m-3 respectivement (Feliú et al., 2020), laissant indiquer une relative stabilité des stocks dans cette zone lors de la période post-bloom à l'automne 2017.
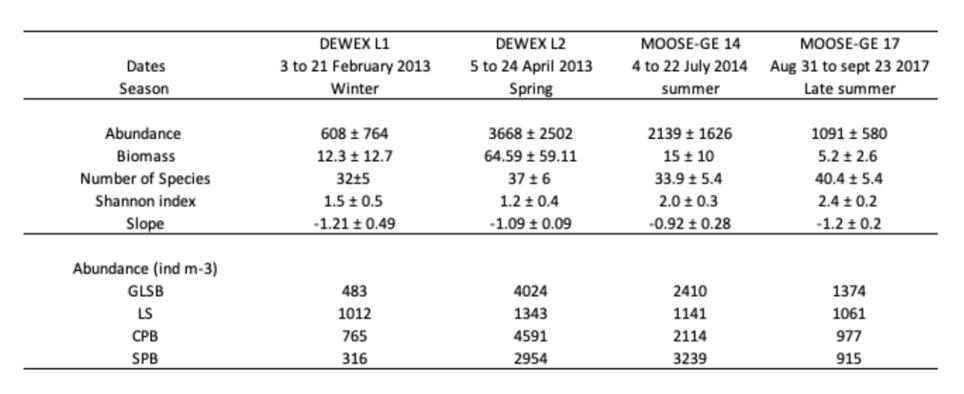
Les résultats antérieurs de diversité, de structure de taille et de fonctionnement du zooplancton en Méditerranée Nord-Occidentale lors de DEWEX 2013 en février (L1) et avril (L2) 2013, et MOOSE-GE en juillet 2014 basées sur des programmes d'échantillonnage similaires (zones échantillonnées et méthodes de prélèvement et d'analyse) permettent aussi de comparer les index principaux du zooplancton. Bien que la présente étude du MOOSE-GE 2017 se soit déroulée plus tard dans la saison estivale (septembre 2017) et lors d'une année avec un processus de convection profonde beaucoup plus faible, il est intéressant de souligner les principales similitudes et différences visibles à partir de ces études (Table 5). La répartition des stations dédiées au filets zooplancton lors de ces campagnes couvrait bien la même zone de Méditerranée Nord-Occidentale, mais les positions le long des radiales ont pu changer d'une campagne à l'autre. Pour effectuer des comparaisons régionales, la zone explorée a été partagée en 4 sous régions : Mer Ligure : LS, Bord du plateau du Golfe du Lion : GLSB, Bassin Provençal central : CPB, Bassin Provençal sud : SPB (Figure 21). Cette subdivision est basée sur des différences environnementales des différentes sous régions mise en évidence par différentes analyses de clusters de variables environnementales (Donoso et al. 2017 ; Feliu 2021).
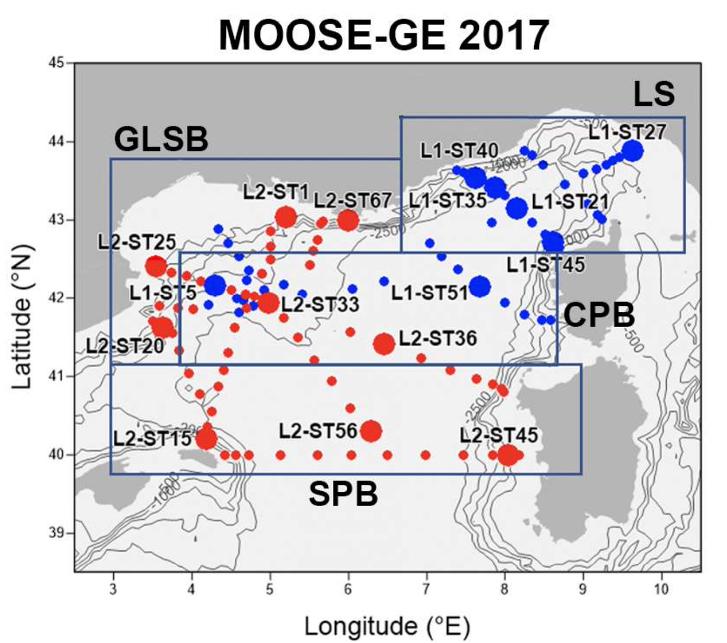
Figure 21. Positionnement des stations de MOOSE-GE 2017 dans les différentes sous-régions : Mer Ligure : LS, Bord du plateau du Golfe du Lion : GLSB, Bassin Provençal central. Les points bleus et rouges correspondent respectivement aux échantillons prélevés pendant les legs 1 et 2 de MOOSE-GE 2017. Les petits points sont les stations avec uniquement des profils CTD et les gros points sont les 16 stations avec des traits de filet à zooplancton en plus des profils CTD. (Thèse G.Feliu , 2021)
Entre les deux campagnes DEWEX, sur l'ensemble de la région, les abondances et biomasses du zooplancton ont été multiplié par 6 et 5 respectivement en l'espace de 2 mois, les stations les plus impactées par le processus d'approfondissement de la MLD (CPB et GLSB) présentant les plus forts contrastes que les zones plus périphériques (SPB et LS; Donoso et al. 2017). A l'augmentation des abondances et biomasse entre l'hiver et le printemps lors de DEWEX (2013) s'associent des baisses de la diversité et des pente NBSS: cela caractérise une augmentation des populations opportunistes à la nouvelle production primaire à la fois en nombre par leur augmentation des taux de reproduction (abondance) et des taux de croissance (biomasse). Ces résultats sont cohérents avec les schémas saisonniers classiques rapportés en Méditerranée (par exemple, Ribera d'Alcalà et al. 2004).
Les abondances observées sur l'ensemble de la région lors des deux campagnes MOOSE, étaient 2 fois moins fortes à MOOSE-GE 2014 par rapport à DEWEX L2, puis à nouveau 2 fois moins fortes entre MOOSE-GE 2014 et 2017 (les facteurs étant encore plus marqués en biomasse). Ce schéma estival global reste vrai à l'échelle des régions CPB et GLSB, mais parait différent pour la zone SPB impactée par des températures plus fortes en été liées aux remontées des eaux atlantiques réchauffées du bassin Algérien repoussant le front Nord-Baléares s'étirant vers le nord, et pour la zone LS, plus individualisée par une circulation moins intense en été (à la fois du courant nord et des structures à mésoéchelle associées). Cette comparaison doit néanmoins garder en mémoire le contexte annuel différent de la situation hydrologique générale à l'échelle de la Méditerranée Nord-Occidentale avec un processus fort de convection profonde durant l'hiver 2013-14 et un processus beaucoup plus faible durant l'hiver 2016-17.
Tableau 5. Comparaison des valeurs moyennes de l'abondance du zooplancton, de la biomasse estimée, du nombre d'espèces, de l'indice de diversité de Shannon, et de la pente NBSS, sur l'ensemble des stations d'échantillonnage de DEWEX 2013, MOOSE-GE 2014 et MOOSE-GE 2017; ainsi que les variations des abondances moyennes par régions (Mer Ligure : LS, Bord du plateau du Golfe du Lion : GLSB, Bassin Provençal central : CPB, Bassin Provençal sud : SPB. NB : Le nombre de stations par région a légèrement différé entre les quatre campagnes. (Thèse G. Feliu , 2021)
L'apport récent de la génomique environnementale dans les campagnes MOOSE-GE depuis 2017 (F. Not, SBR Roscoff, M. Lescot MIO Marseille) a pour objectif de : 1) décrire la diversité biologique des assemblages de plancton dans un contexte océanographique, 2) mettre en ?uvre une procédure d'échantillonnage simple et standardisée à inclure dans la surveillance à long terme de la zone, 3) diffuser rapidement des données significatives à la communauté. Les observations se font à l'aide de filets à plancton et de prélèvement d'eau de mer (bouteilles Niskin). Ces derniers permettent en particulier d'étudier la diversité microbienne afin de: 1) décrire la diversité moléculaire par méta-barcoding des assemblages planctoniques à travers 2 fractions de taille (bactéries/picoeucaryotes et phyto/petit zoo), 2) avoir accès aux fonctions biologiques des procaryotes par le biais d'analyses de méta-génomique. Les observations par filets permettent : 1) de décrire la diversité moléculaire des composants zooplanctoniques (>64 ?m, >200 ?m, > 500 ?m), 2) de quantifier le mésozooplancton grâce à l'imagerie automatisée in-situ. A chaque campagne MOOSE-GE, un total de 400 échantillons pour les analyses génomiques est collecté.
Des premiers échantillons provenant de filets et de bouteilles Niskin des trois années (de 2017 à 2019) ont été séquencés par méta-barcoding en ciblant l'ADNr 18S (fragment V4). Ces séquences permettent d'identifier la diversité du plancton eucaryote. Au total, 271 échantillons de 2017 et 2018 ont été analysés et intégrés dans le serveur « The Ocean Barcode Atlas » (OBA ; Vernette et al. 2021) ainsi que les données environnementales (physiques et chimiques) associées. Les échantillons de méta-barcodes de 2019 sont en cours d'intégration dans la base de données.
Le serveur OBA permet d'obtenir différentes analyses des données de méta-barcoding comme la biogéographie de séquences 18S associées à un taxon, la diversité de différents sous groupes mais également des analyses d'alpha- et beta-diversité. Ce serveur web en en cours d'évolution pour intégrer les données temporelles des différentes campagnes MOOSE-GE. Cependant des résultats préliminaires sont présentés ici. Des tendances sur ces deux années sont observées en termes d'évolution de la diversité des différents super-groupes d'eucaryotes (Figure 22). En 2018, une augmentation de l'abondance des Rhizaria est observée dans les filets, les straménopiles quant à eux sont plus abondants dans les échantillons des bouteilles Niskin en 2018 avec une apparition en Deep (ou mésopélagique).
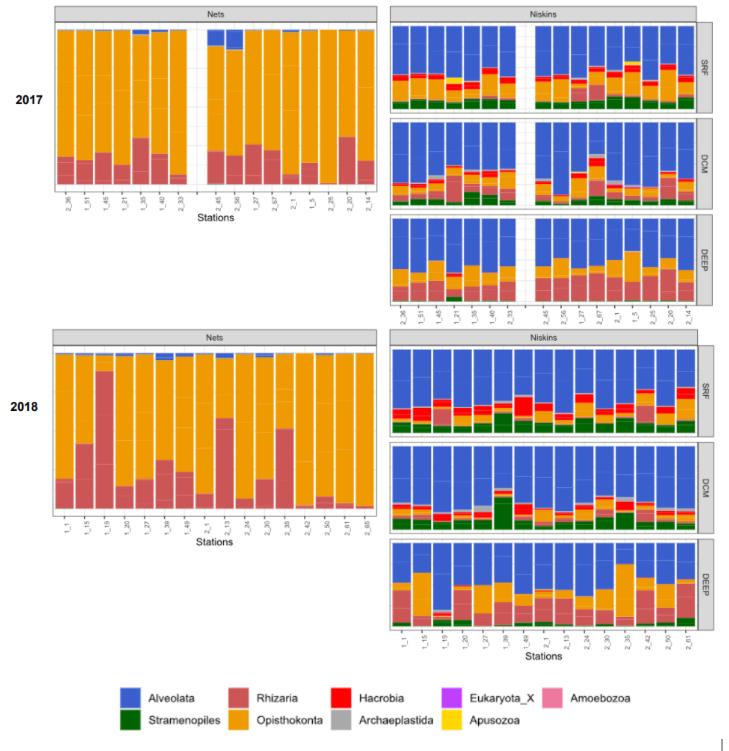
Figure 22: Abondances des différents super groupes d'eucaryotes en 2017 et 2018
A titre d'exemple et de façon plus spécifique, dans la figure 23 ci-dessous, la biogéographie de l'abondance du groupe des radiolaires est représentée à partir des les bouteilles Niskin échantillonnées en surface, DCM ou Deep (ou mésopélagique) et dans les filets. On peut remarquer que l'abondance relative de ce groupe taxonomique est plus importante dans les échantillons Deep des bouteilles Niskin et dans les filets en 2018.
Ce type d'analyses de biogéographie pour différents rangs taxonomiques en lien avec les paramètres environnementaux peut-être étendu et réalisé facilement pour les différents groupes d'intérêts via le site OBA (https://oba.mio.osupytheas.fr/ocean-atlas). Les analyses de diversité procaryote et de méta-génomique (plus coûteuse en ce qui concerne la production et l'analyse des données) sont en phase de préparation.
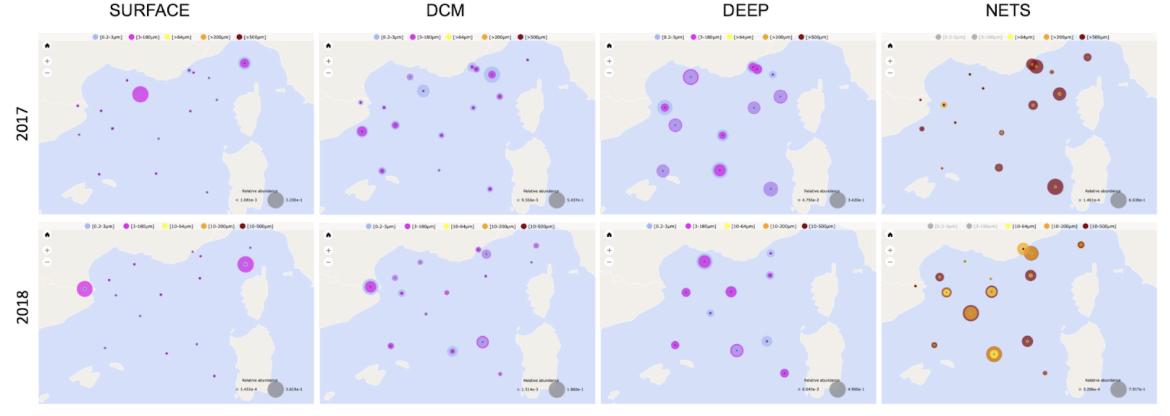
Figure 23 : Biogéographies des radiolaires dans les échantillons de bouteilles Niskin et les filets
- Développement d'un réseau de neurones en Méditerranée entraîné par les données des campagnes MOOSE-GE
Les données biogéochimiques des campagnes MOOSE-GE de 2010 à 2016 ont été utilisées dans le cadre du développement de méthodes neuronales pour l'océanographie. Ces données qualifiées ont été regroupées afin de former un jeu de données de 35 campagnes en mer et une série temporelle entre 1976 et 2018 (Fig.24) présentant des mesures bouteilles de la surface au fond d'O2, température, salinité ainsi que macronutriments (nitrates, phosphates, silicates) et variables du système des carbonates (AT, CT, pH). Ce jeu de deu de données a été séparé en 2 sous-parties, un jeu d'entraînement et un jeu de validation, et a permis l'entraînement de méthodes de machine learning (réseaux de neurones artificiels) permettant de dériver des variables biogéochimiques à partir de variables systématiquement mesurées. Cette méthode s'appelle CANYON-MED pour ¿CArbonate system and Nutrients concentration from hYdrological properties and Oxygen using a Neural network in the MEDiterranean Sea¿ (Fourrier et al., 2020). Ainsi, à partir de la position dans le temps et l'espace ainsi que les température, salinité et O2, il est possible en appliquant la méthode CANYON-MED de dériver les concentrations en nutriments et les variables du système des carbonates avec une précision donnée.
Ces méthodes ont un large champ d'application, par exemple sur les plateformes autonomes et mouillages faisant l'acquisition de mesures d'O2 mais manquant encore cruellement de capteurs biogéochimiques. De plus, l'utilisation de ces méthodes neuronales peut permettre de détecter des valeurs anormales qui n'auraient pas été qualifiées auparavant en comparant les estimations de CANYON-MED aux données mesurées in situ.
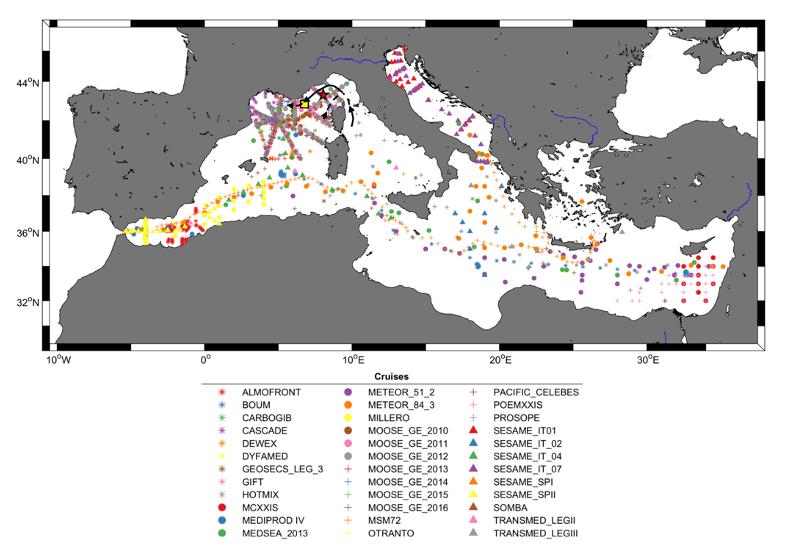
Figure 24 : Localisation des 35 campagnes en mer (dont les campagnes MOOSE-GE 2010 à 2016), et des séries temporelles DYFAMED (étoile rouge) et ANTARES (carré jaune) en Méditerranée utilisées pour entrainer la méthode neuronale CANYON-MED (tiré de Fourrier et al., 2020).
Par conséquent, en appliquant CANYON-MED sur le large réseau de flotteurs Argo équipés d'O2 (Figure 25), les variables biogéochimiques peuvent être dérivées et leurs variations temporelles étudiées, par exemple en réponse à la convection dans un contexte de réduction des événements de formation d'eau dense (Somot et al. 2006 ; Pagès et al. 2020 ; Soto-Navarro et al. 2020). Dans le Golfe du Lion, cette application a permis de déterminer que les nitrates augmentent dans les couches intermédiaires et tous les nutriments augmentent en profondeur, alors que dans la Mer Ligure, seuls les nitrates et les silicates en profondeur augmentent sur la période d'étude (Figure 26). Cependant, ce découplage entre les nitrates et les phosphates pourrait conduire à une évolution des rapports molaires des nutriments par rapport au modèle de Redfield dans les années à venir. De plus, les séries temporelles long terme d'acidification font toujours défaut pour la mer Méditerranée. L'approche novatrice utilisant le pH dérivé de CANYON-MED a permis d'estimer l'acidification dans différentes couches de la colonne d'eau. Dans l'ensemble pour les variables du système des carbonates, le CT augmente avec une diminution simultanée du pH et a été fortement affecté par les événements de mélange. Cette évolution est visible dans toute la colonne d'eau, ce qui pourrait affecter l'écosystème marin profond dans un avenir proche.
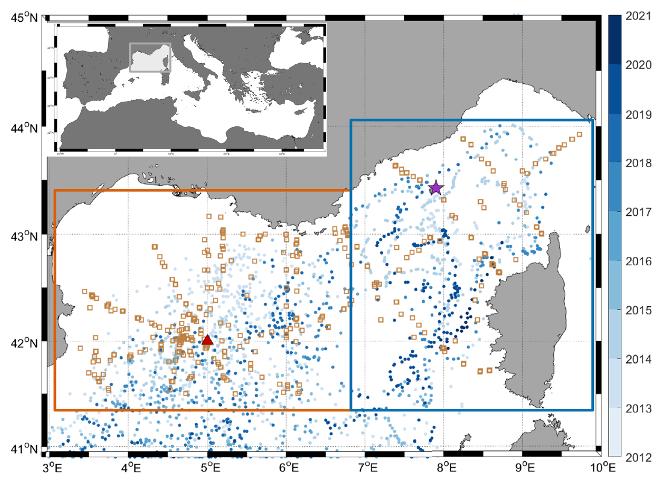
Figure 25 : Position des flotteurs profileurs Argo équipés de capteurs O2 en fonction de la date du profil (cercles bleus), des stations de campagnes en mer (carrés bruns), et des mouillages LION (triangle rouge) et DYFAMED (étoile violette) dans le Golfe du Lion (boîte orange) et la Mer Ligure (boîte bleue) respectivement. Deux régions sont distinguées : le Golfe du Lion et la Mer Ligure (encadré orange et bleu respectivement) (Fourrier et al, in review).
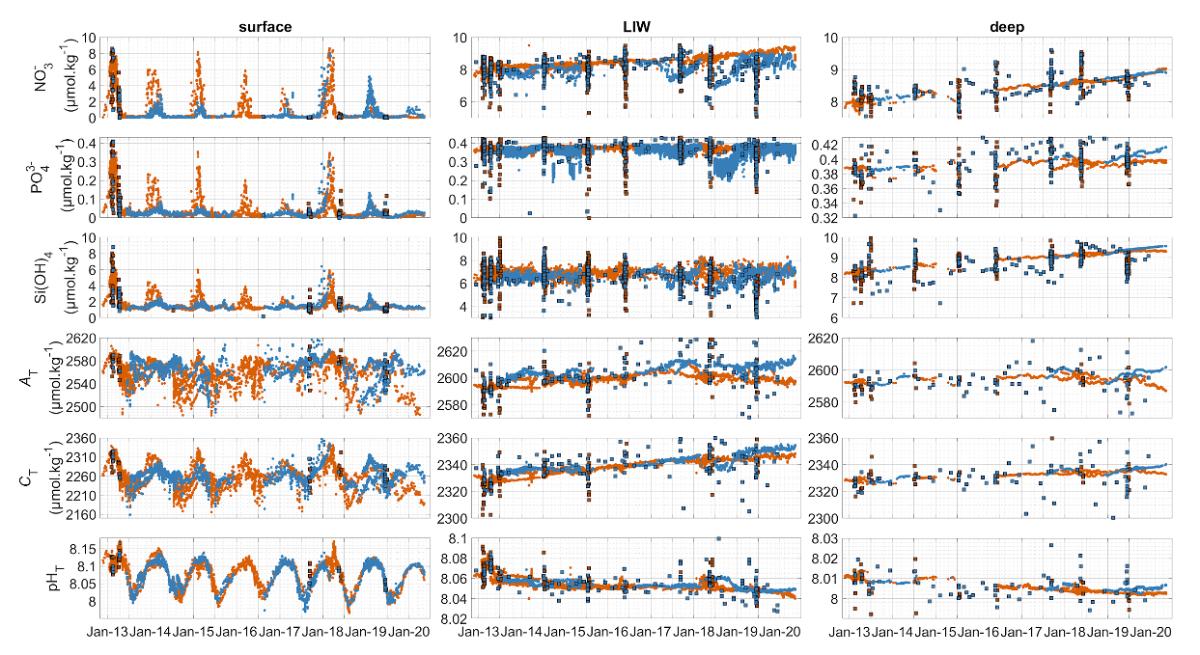
Figure 26 : Évolution biogéochimique de la mer Méditerranée Nord-Occidentale. Les panneaux a-f montrent l'évolution dans les eaux de surface (avec une grande variabilité saisonnière), les panneaux g-l dans les eaux intermédiaires (à la profondeur de la LIW), et les panneaux m-r à 2000 m, pour les nitrates (a, g, m), les phosphates (b, h, n), les silicates (c, i, o), l'alcalinité totale (d, j, p), le carbone total (e, k, q) et le pHT (f, l, r). Les valeurs dérivées de CANYON-MED à partir des données des flotteurs Argo sont représentées par des points (orange pour le Golfe du Lion et bleu pour la Mer Ligure). Les mesures des bouteilles (données des campagnes MOOSE-GE) sont superposées sous forme de carrés colorés en fonction de leur zone. Les données des bouteilles proviennent d'une correspondance de 15 jours - 25 km - avec les valeurs dérivées CANYON-MED des flotteurs Argo (Fourrier et al, in review).
Le déploiement et la récupération de plateformes autonomes dans le cadre des campagnes MOOSE-GE contribue à un effort constant tout au long de l'année réalisé avec des moyens de façade (déploiements et/ou récupérations de gliders et de flotteurs avec zodiac ou lors des campagnes de suivi mensuel). En revanche, c'est la seule campagne qui visite l'ensemble du bassin et permet des opérations de récupération de gliders ou de flotteurs profileurs lorsqu'ils sont loin des côtes françaises (> 20 milles nautiques) en dehors de campagnes d'opportunité. Elle est donc essentielle à cet effort d'observation multi- échelles. De plus, les données de la campagne MOOSE-GE sont utilisées comme points de référence pour l'ensemble des observations réalisées à partir de gliders et de flotteurs profileurs (T, S, courants, Chl-a, O2, turbidité, nitrate). Après un travail important de calibration, la calibration des données récoltées par le système dans son ensemble permet de qualifier les variations à l'échelle du bassin avec une bonne précision et un bon niveau de significativité.
Entre les zones côtières et le large, on trouve des contrastes hydrographiques entre les différentes masses d'eau, en particulier en surface et à profondeur intermédiaire. En effet, la circulation de bord en équilibre géostrophique est contrôlée par les contrastes de densité forts au niveau du talus. Ainsi, dans les trois zones côtières de la Figure 27, on s'attend à trouver des Eaux Atlantiques moins salées en s'approchant de la côte et des Eaux Levantines Intermédiaires plus marquées (i.e. plus chaudes et plus salées).
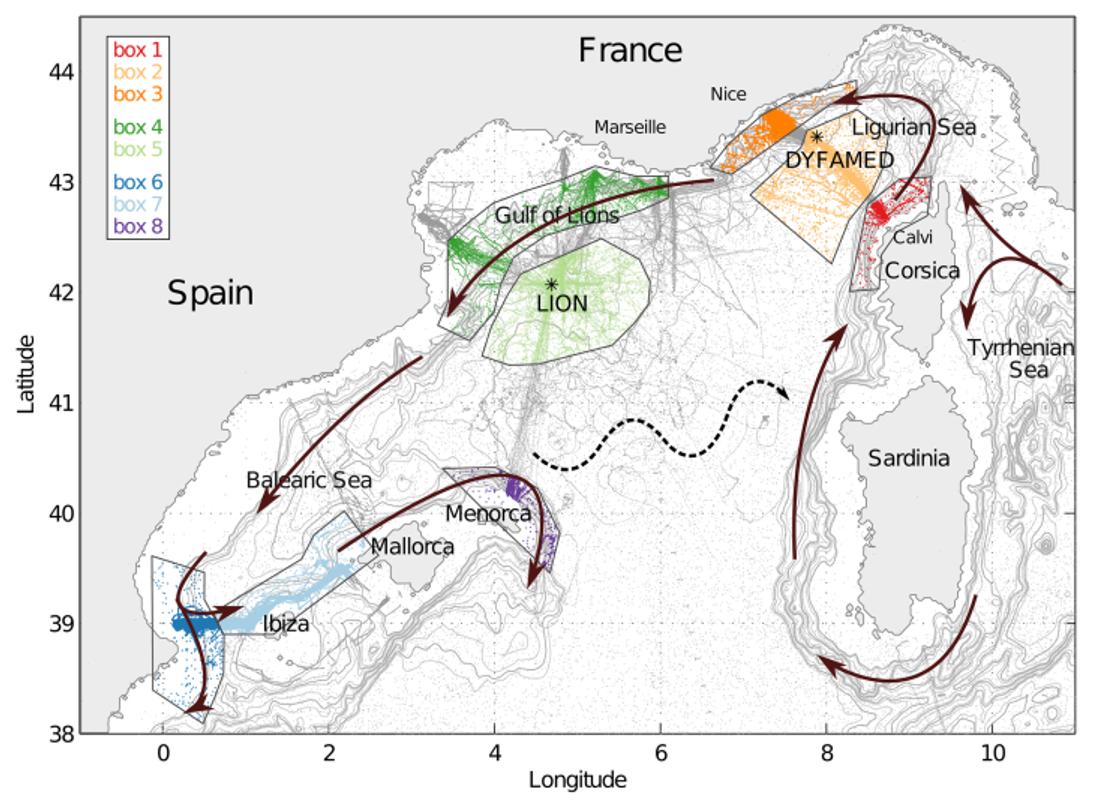
Figure 27 : Ensemble des observations de profils CTD réalisés pour sur la période 2007-2018 incluant les campagnes MOOSE-GE, ainsi que flotteurs profileurs et gliders déployés durant la campagne. Domaines côtier/talus en couleurs foncée et hauturiers en pastel. Zones d'observation clés : délimité en vert, on trouve la zone du Golfe du Lion (à l'Ouest) et en orange la Mer Ligure (à l'Est), en bleu la zone des Baléares (tiré de Margirier et al 2020).
Une approche statistique basée sur ces boîtes et sur une détection automatique par profil des caractéristiques des masses d'eau mesurées sur la base des propriétés T-S de chaque profil, permet de construire des index de température et salinité qui rendent compte de ces contrastes et de la variabilité des propriétés des masses d'eau. On utilise le signal qui correspond aux 10% des valeurs extrêmes (minimales ou maximales selon la masse d'eau et la profondeur) dans un intervalle de temps donné (typiquement 10 jours).
On distingue :
- les Eaux Atlantiques sont définies par la température et la salinité moyenne au-dessus de l'isopycne 28.95 kg/m3 ;
- les Eaux Levantines Intermédiaires sont définies par la température et la salinité moyenne en dessous de l'isopycne 29 kg/m3 et au-dessus de 29.1 kg/m3 ouùse trouvent les maxima de température et de salinité typiques de cette masse d'eau;
- les Eaux Profondes de la Méditerranée sont suivies par le signal moyen de température et de salinité mesuré en dessous de 1000 m.
Avec ces considérations il est possible d'extraire des index de variabilité significatifs (Figures 28, 29 et 30) qu'il faudra comparer au forçage atmosphérique et aux estimations de transports à différent niveaux (estimations des transports au canal de Corse et de Sardaigne-Tunisie, de Sicile en collaboration avec l'Italie). Ces index, que l'on peut extraire de simulations numériques d'une manière similaire, constituent un défi pour la modélisation. Le système d'observation MOOSE calibré par la campagne MOOSE-GE est opérationnel depuis 8 ans et des analyses sur l'interannuel sont désormais possibles. Ces index fournissent de l'information sur l'amplitude du signal recherché dans les données et la précision des mesures récoltées pendant la campagne et des mesures flotteurs, gliders, mouillages, etc. qui sont calibrées par les données des campagnes MOOSE-GE est bien en adéquation.
- Évolution des Eaux Atlantiques de surface
La Figure 28 montre l'évolution des caractéristiques des Eaux Atlantiques. Elles varient principalement de manière saisonnière pour la température avec une amplitude du cycle annuel d'environ 4 à 5°C. En revanche, la salinité varie elle sur des échelles de temps plus longues avec une amplitude d'environ 0.1. On remarque qu'en 2011, toutes les zones (au large et côtières) étaient caractérisées par des faibles valeurs de salinité. L'évaporation estivale peut également favoriser l'augmentation de salinité comme elle semble augmenter en phase avec la température lors des été 2010 et 2011 en mer Ligure.
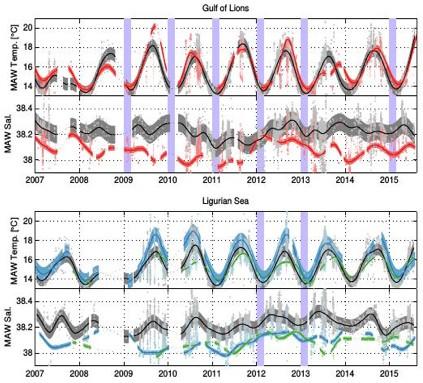
Figure 28 : Évolution de la température et salinité des Eaux Atlantiques sur la période 2008-2015 dans le Golfe du Lion (en haut) et en mer Ligure (en bas). Les points gris représentent les observations faites dans les zones au large et en couleur dans les zones côtières décrites sur la Figure 27. Les courbes épaisses représentent le signal moyen lissé sur une fenêtre de 90 jours pour rendre compte des variations saisonnières et les zones colorées représentent la déviation standard des données. Pour construire la courbe épaisse des zones côtières, on a uniquement gardé 10% des plus petites valeurs de salinité et des plus grandes valeurs de température dans chaque intervalle de 10 jours. Les mois d'hiver (JFM) caractérisées par de forts épisodes de convection sont indiqués en violet.
En hiver, on observe des pics locaux qui suivent de 1-2 mois les épisodes de convection profonde. En effet, le mélange hivernal et les processus de restratification de la zone de mélange hivernal, ramènent en surface du sel provenant des couches profondes et intermédiaires plus salées (S > 38.5). Ces phénomènes se conjuguent avec des phénomènes d'évaporation pour dominer l'apport d'eaux Atlantiques moins salées (~35 en salinité à Gibraltar). Ce phénomène semble être plus marqué en Mer Ligure, probablement parce que la zone mélangée contient plus de sel. Le mélange hivernal y est moins profond et les Eaux Levantines Intermédiaires plus salées, plus proches de leurs sources dans le bassin, le Canal de Corse et le Sud de la Sardaigne. La salinité a augmenté de 0.05-0.1 dans la couche d'Eaux Atlantiques lors du printemps qui suit les épisodes de convection des hivers 2012 et 2013. Les Eaux Atlantiques ne sont plus détectées lorsque la densité de surface excède 1028.95 kg m-3, ce qui provoque une discontinuité dans les séries temporelles. L'amplitude de variation de la salinité des Eaux Atlantique est de l'ordre de 0.1 ce qui peut avoir une influence importante sur le phénomène de formation d'eaux intermédiaires et profondes.
- Évolution des Eaux Levantines Intermédiaires et des Eaux Profondes
Les Eaux Levantines Intermédiaires jouent un rôle très important dans le processus de renouvellement des Eaux Profondes de la Méditerranée Occidentale. En particulier, un excès de chaleur et de sel à profondeur intermédiaire combiné à des conditions atmosphériques favorables ont provoqué un changement abrupt des propriétés des Eaux Profondes en 2005 (Western Mediterranean Transient, WMT). Les couches intermédiaires constituent un lieu d'accumulation privilégié de chaleur et de sel avant d'être transféré par mélange vertical dans les couches profondes de l'océan (Fig. 29)
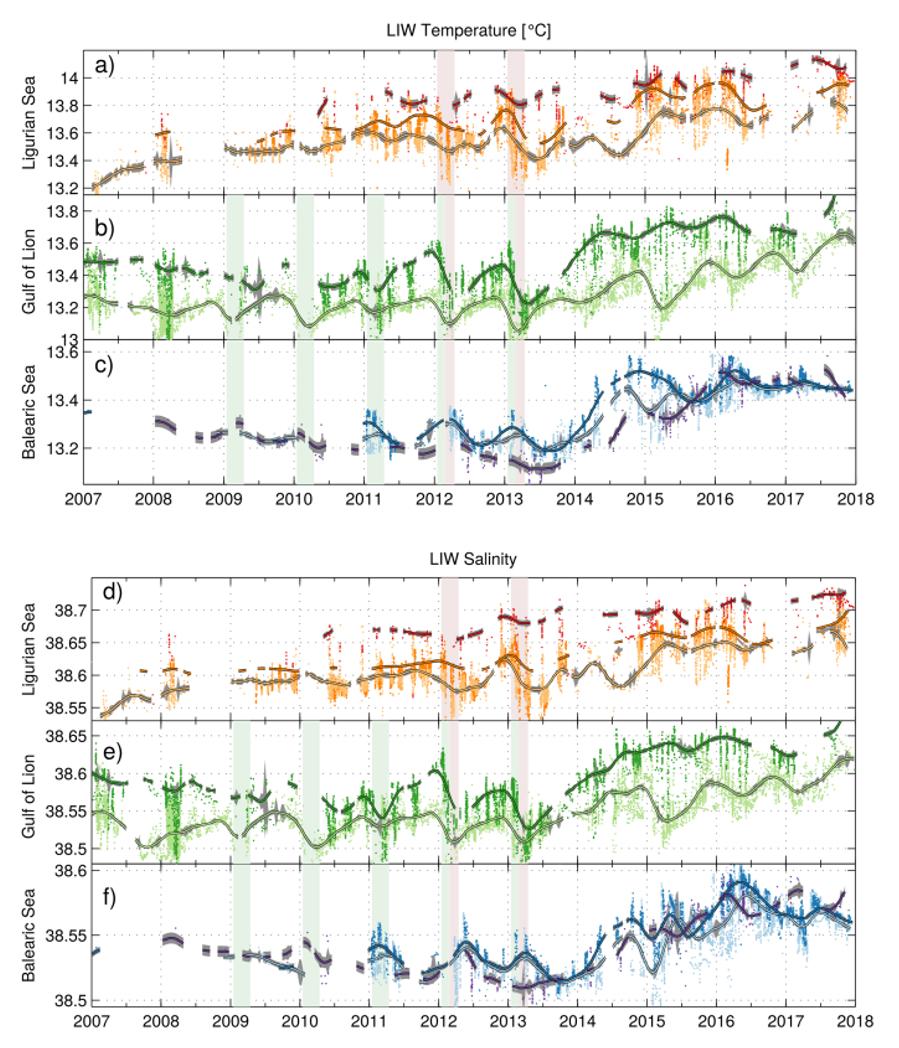
Figure 29 : Évolution de la température et salinité des LIW sur la période 2008-2018 dans le Golfe du Lion (en vert), en mer Ligure (en orange) et mer des Baléares (en bleu). Les courbes en gras représente le signal moyen lissé sur une fenêtre de 90 jours pour rendre compte des variations saisonnières, et la zone colorée autour de la courbe représente la déviation standard des données. Les mois d'hiver (JFM) caractérisés par de fort épisodes de convection sont indiqués en violet.
Les épisodes de convection profondes des 5 hivers consécutifs (2009 à 2013) et celui de 2015 sont facilement identifiables dans le Golfe du Lion. Lors de chaque épisode, on constate une diminution de la température (0.1-0.3°C) et de la salinité (0.02-0.05) des Eaux Levantines Intermédiaires, qui traduit le transfert de chaleur et de sel vers les couches profondes (voir figure 18). Ce transfert de chaleur et de salinité est également observé lors des hivers 2012 et 2013 en Mer Ligure, où la couche de mélange a atteint 1000 m environ. On voit d'ailleurs qu'au même moment, la température et la salinité des Eaux Profondes de la Méditerranée Occidentale augmentent de respectivement 0.01-0.02°C et 0.01.
Le signal de salinité des couches profondes montre bien l'augmentation temporaire de la salinité des Eaux Profondes au moment de leur renouvellement (voir figure 30). Sous l'action d'échanges latéraux entre les Eaux Profondes nouvellement formées et celles situées en dehors de la zone de convection qui ont conservées des propriétés moins salées, le signal décroit ensuite régulièrement jusqu'à l'hiver suivant.
L'augmentation de salinité (resp. de température) atteint 0.015 (resp. 0.025°C) entre 2009 et 2015 soit un rythme de 0.0025 an-1 (resp. 0.004°C an-1) comparable avec l'estimation récente de Marty and Chiavérini, (2010) en Mer Ligure entre 1995 et 2005. Ce rythme d'augmentation de la température et de la salinité des Eaux Profondes de la Méditerranée Occidentale reste quand même plus important que ceux mesurés entre les années 1960/70 et 1990, ce qui suggérerait une amplification du phénomène, en lien avec le réchauffement climatique et les changements récents du climat Méditerranéen.
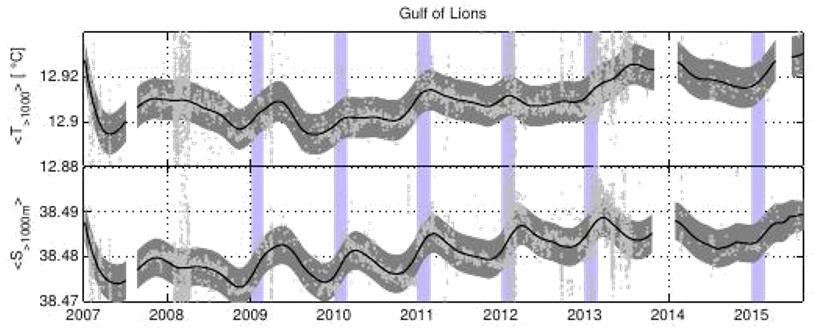
Figure 30 : Évolution de la température et salinité des WMDW sur la période 2008-2015 dans le Golfe du Lion. Les observations de température moyenne en dessous de 1000 m sont représentées en gris, la courbe noire est le signal moyen lissé sur une fenêtre de 90 jours pour rendre compte des variations saisonnières, et la zone grisée représente la déviation standard des données. Les mois d'hiver (JFM) caractérisées par de fort épisode de convection sont indiqués en violet.
- Contraste Est/Ouest et côte/large
Les caractéristiques des LIW est en augmentation globale sur notre période d'étude, mais l'augmentation est la plus notable lors de l'année 2014, qui est caractérisée par une faible convection dans tout le bassin. On observe une augmentation de la température (+0.3°C dans le Courant Nord et +0.2°C au large) et de la salinité (+0.07 dans le Courant Nord et +0.05 au large) des LIW dans la zone du Golfe du Lion. Avec une température d'environ 13.7°C et une salinité de 38.63 à la fin de la période d'étude, les caractéristiques des LIW du Courant Nord dans le Golfe du Lion se rapprochent de celles de la Mer Ligure. En Mer Ligure, on observe également une augmentation plus étalée et d'amplitude moindre +0.2°C en température et +0.05 en salinité. Comme lors de l'hiver 2014, la convection n'a pas été active dans le bassin, on observe une accumulation de chaleur et de sel particulièrement marquée dans le Golfe du Lion en provenance de la Mer Ligure. Il faut noter qu'en attente d'une calibration finale et solide des données de 2015, celle -ci se base uniquement sur la méthode de la droite de mélange déterminée avec les profils de la sonde CTD qui a pu seulement être calibrée « constructeur » avant la campagne avec une précision de 0.003, que nous vérifierons à posteriori avec une calibration constructeur post-campagne et les analyses des échantillons d'eau. De légers ajustements seront donc probablement faits à ces données dans un futur très proche, mais on peut tout de même parler d'augmentation significative et spectaculaire vu l'amplitude et la rapidité de ces changements.
Enfin, il y a moins de gradient côte/large en Mer Ligure. Cela est certainement lié à la taille restreinte du bassin dans cette zone et à la présence de deux courants de bord (le Courant Ouest-Corse et la Courant Nord) qui génèrent des flux latéraux de chaleur et de sel par instabilité hydrodynamique. Les épisodes hivernaux de mélange franchissant la couche de LIW sont également moins fréquents que dans le Golfe du Lion (2 épisodes contre 6 ou 7 lors des 8 hivers de la période 2008/2015, voir figure 29) ce qui limite la redistribution verticale de chaleur et de sel sur la colonne d'eau et facilite son accumulation. L'absence de convection profonde dans le Golfe du Lion en 2014 et l'augmentation des caractéristiques T/S des LIW qui en a suivi suggère une propagation rapide (~1 an) des masses d'eau intermédiaires de l'est à l'ouest du bassin.
- Circulation à mésoéchelle
En plus de renseigner les grands traits de la circulation du bassin, les campagnes MOOSE-GE permettent également d'observer, voire même de quantifier la dynamique de tourbillons rencontrés (Figure 31). Le plan de campagne étant prédéfini, les observations de tourbillons de méso-échelle durant MOOSE-GE sont fortuites, mais toutefois régulières (en moyenne 1-3 tourbillon par campagne). Ces tourbillons sont extrêmement difficiles à détecter du fait de leur petite taille (rayon de 5-10km, vitesses de rotation 0.1-0.2 m/s). C'est uniquement du fait d'une population relativement importante (une dizaine sur le bassin) et du nombre de stations conséquent effectué durant les campagnes MOOSE-GE que l'on a la chance de pouvoir étudier chaque années de nouveaux spécimens.
Ces tourbillons peuvent ont généralement deux origines différentes (Bosse et al, 2015, 2016):
- soit ils se forment durant l'hiver après la phase de mélange profond par restratification. Ils se repèrent en général par une température/salinité quasi constante sur la verticale et un contenu en oxygène dissous plus important.
- soit ils se forment par détachement et instabilité de la circulation de bord, avec un signal fort sur les LIW, qui sont plus marquées que le signal habituel du large.
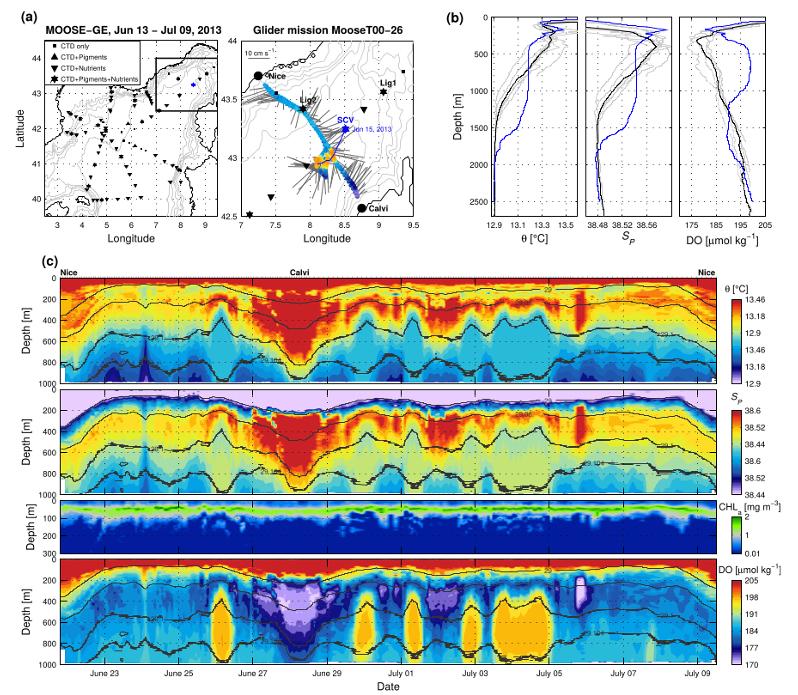
Figure 31 : Exemple du tourbillon échantillonné de manière concomitante pendant la campagne MOOSE-GE 2013 et par un glider MOOSE déployé sur la radiale T00 Nice-Calvi (tiré de Bosse et al 2017). (a) carte des stations réalisées pendant MOOSE-GE 2013 avec paramètres échantillonnés, et trajectoire du glider; (b) Profils de température, salinité et oxygène réalisés pendant la campagne. c) Température potentielle, salinité, fluorescence et oxygène dissous observés par le glider dans le tourbillon.
Un exemple marquant a été le tourbillon rencontré en 2013 durant la campagne MOOSE-GE entre le France et la Corse. Un glider MOOSE déployé sur la radiale T00 à la même période a permis d'en faire une caractérisation et un suivi fin durant près d'une semaine (Figure 31). Les données récoltées pendant la campagne ont permis d'étendre énormément le champ d'analyse en caractérisant ce tourbillon sur les variables biogéochimiques, et les pigments du phytoplancton (Bosse et al, 2017).
Les données de courantologie acquises sur le bateau par S-ADCP et L-ADCP (ship-ADCP et lowered-ADCP) permettent également de compléter la vision lacunaire des profils CTD lorsqu'un tourbillon est rencontré (Figure 32). Cela permet notamment de procéder à l'analyse de leur dynamique, en détectant leur centre et en les caractérisant en termes de maximum de vitesse, etc. Ces mesures de vitesses sont cruciales, et une fois combinées avec les données CTD, ainsi que toutes les analyses biogéochimiques et biologiques faites à bord, les impacts de ces circulations à (sous)méso-échelle pourront être évalués de manière plus systématique. Cela permettra notamment de mieux comprendre leur rôle dans les équilibres physiques, biogéochimiques et biologiques du bassin Méditerranéen. L'évolution de leurs propriétés au cours de la série de campagne MOOSE-GE nous permet également de mieux appréhender le réchauffement et la salinisation rapides des eaux de la Méditerranée en réponse au changement climatique.
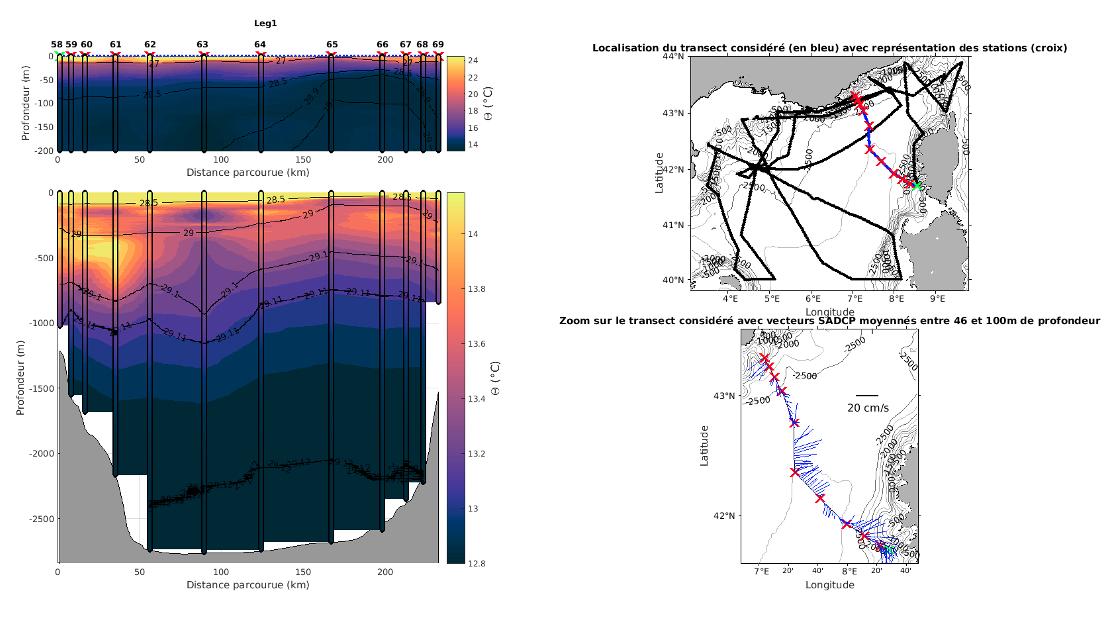
Figure 32 : Exemple d'une section de température (gauche) et courants mesurés par S-ADCP (droite) durant la campagne MOOSE-GE 2021 montrant la présence d'un tourbillon anticyclonique profond au niveau de la station 63 (stage M2 E. Chevillon, 2022).
- Le réseau de flotteurs Argo-O2
En appui au réseau MOOSE, la composante Argo-O2, initiée durant la SOP2 HYMEX (hiver 2012/2013), a permis de mieux contraindre la dynamique de l'oxygène de part et d'autre du bassin (projet GMMC MOOXY). Ce réseau est dimensionné aujourd'hui de manière opérationnelle pour le suivi à long terme des masses d'eau. Depuis fin 2012, 21 flotteurs Argo-O2 ont été déployés lors des campagnes MOOSE-GE, en plus du réseau de flotteurs NAOS (flotteurs BGC-Argo). Cela correspond environ à un ensemencement de 3-4 flotteurs tous les 2 ans. Actuellement, 4 flotteurs sont toujours actifs (Fig. 33)
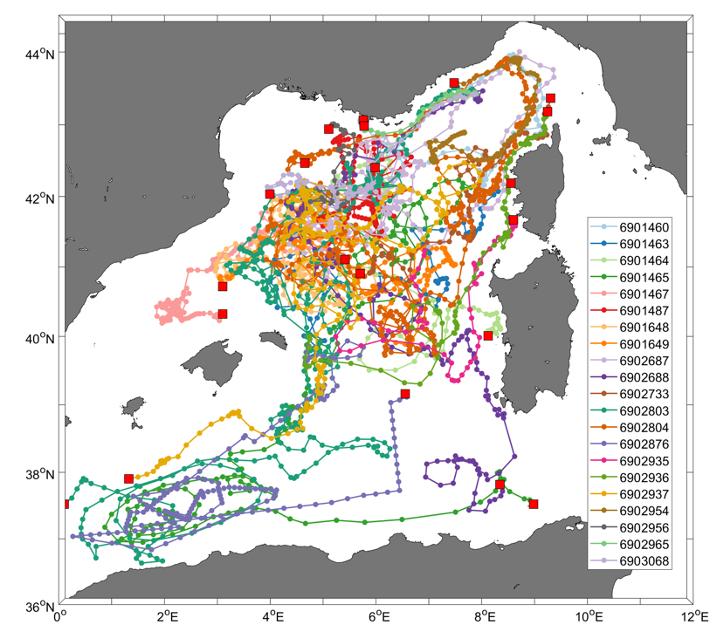
Figure 33 : Réseau de flotteurs Argo-O2 dans le bassin Occidental déployés pendant les campagnes MOOSE-GE. Le carré rouge représente la dernière position des flotteurs.
Les déploiements se font lors des campagnes MOOSE-GE, ce qui permet d'avoir plusieurs profils CTD-O2 de référence, corrigés avec les données in situ Winkler et, in fine, d'obtenir des données O2 de flotteurs Argo ajustées en temps différé. Ces données ajustées sont envoyées régulièrement au GDAC (Global Data Assembly Center) CORIOLIS. Les corrections de ces données se font en appliquant les méthodes de correction de Takeshita et al. (2013) et Bittig et Kortzinger (2015). Ces méthodes et les procédures d'ajustement suivent la dernière version du « cookbook » de Coriolis (Thierry et al., 2021).
Les premières données ont permis de tracer l'impact de la convection hivernale sur la ventilation des masses d'eau intermédiaires et profondes (Fig.34), d'estimer un volume d'eau dense ventilée qui aura un fort impact sur les écosystèmes marins profonds (Coppola et al., 2017) et également de suivre l'expansion de la couche de minimum d'O2 dans les eaux intermédiaires qui joue un rôle clef dans l'export de matière organique (export vs. reminéralisation).
Les déploiements se font lors des campagnes MOOSE-GE, ce qui permet d'avoir plusieurs profils CTD-O2 de référence, corrigés avec les données in situ Winkler et, in fine, d'obtenir des données O2 de flotteurs Argo ajustées en temps différé. Ces données ajustées sont envoyées régulièrement au GDAC (Global Data Assembly Center) CORIOLIS. Les corrections de ces données se font en appliquant les méthodes de correction de Takeshita et al. (2013) et Bittig et Kortzinger (2015). Ces méthodes et les procédures d'ajustement suivent la dernière version du « cookbook » de Coriolis (Thierry et al., 2021).
Les premières données ont permis de tracer l'impact de la convection hivernale sur la ventilation des masses d'eau intermédiaires et profondes, d'estimer un volume d'eau dense ventilée qui aura un fort impact sur les écosystèmes marins profonds (Coppola et al., 2017) et également de suivre l'expansion de la couche de minimum d'O2 dans les eaux intermédiaires qui joue un rôle clef dans l'export de matière organique (export vs. reminéralisation).
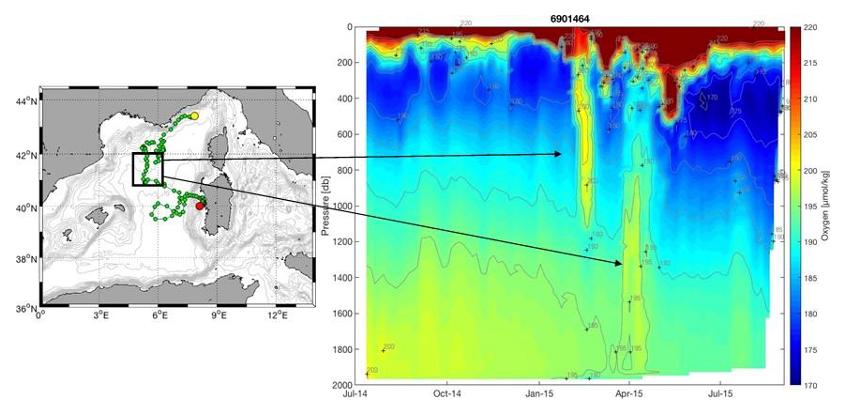
Figure 34 : Données O2 acquises par le flotteur Argo 6901464 de juillet 2014 (point jaune) à juillet 2015 (point rouge). Le signal du mélange hivernal est bien marqué par les concentrations en O2 plus élevées jusqu'à 1000m à l'approche du centre du Golfe du Lion en mars 2015 puis par une ventilation d'O2 en profondeur bien visible en avril 2015. Par la suite, la couche du minimum d'O2 apparait dans les eaux intermédiaires et s'intensifie au niveau de la Sardaigne là où le signal de la LIW est plus marqué.
En intégrant le large nombre de flotteurs, il est possible de reconstruire la dynamique de l'O2 et d'ainsi visualiser l'impact des évènements de mélange hivernal profond sur la dynamique du minimum d'O2 dans les eaux intermédiaires et sur l'ensemble de la colonne d'eau (Figure 35).
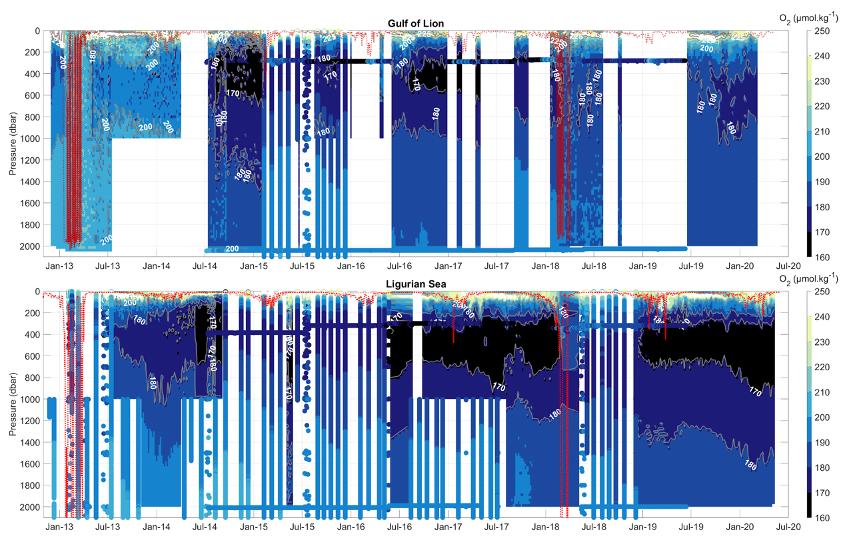
Figure 35 : Séries temporelles d'O2 issu de flotteurs Argo, de mouillages et de campagnes MOOSE-GE dans le Golfe du Lion (a) et en mer Ligure (b) et profondeur de la couche de mélange (ligne rouge). Les approfondissements hivernaux de la couche de mélange détruisent le minimum d'O2 présent dans les eaux intermédiaires (LIW) et celui-ci se reconstruit avec une vitesse, une épaisseur et une concentration entre les 2 zones (Fourrier et al., in review).
Les données des flotteurs O2 permettent également, une fois l'O2 corrigé, de servir de référence pour évaluer des réanalyses de modèles biogéochimiques adaptés à la Méditerranée. La figure 36 montre le cas d'une réanalyse sur la période 1999-2019 à une résolution 1/24° développée dans le cadre du Copernicus Marine Environment Monitoring Service (CMEMS).
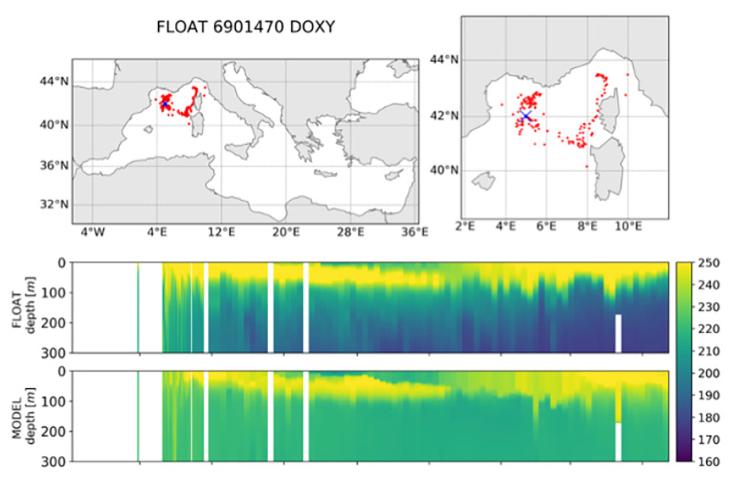
Figure 36 : Données O2 acquises par le flotteur Argo 6901470 de mars 2013 (croix bleue) à mai 2014. Les panneaux montrent la concentration en O2 issue du flotteur et d'un modèle biogéochimique (tiré de Cossarini et al., 2021).
Références non listées dans la bibliographie des campagnes MOOSE-GE:
Articles avec comité de lecture (ACL):
Belgacem, M., Schroeder, K., Barth, A., Troupin, C., Pavoni, B., Raimbault, P., Garcia, N., Borghini, M., and Chiggiato, J.: Climatological distribution of dissolved inorganic nutrients in the western Mediterranean Sea (1981¿2017), Earth Syst. Sci. Data, 13, 5915¿5949, https://doi.org/10.5194/essd-13-5915-2021, 2021.
Feliu, G., Pagano, M., Hidalgo, P., Carlotti, F., 2020. Structure and function of epipelagic mesozooplankton and their response to dust deposition events during the spring PEACETIME cruise in the Mediterranean Sea. Biogeosciences 17, 5417¿5441. https://doi.org/10.5194/bg-17-5417-2020.
Nowaczyk, A., Carlotti, F., Thibault-Botha, D., Pagano, M., 2011. Distribution of epipelagic metazooplankton across the Mediterranean Sea during the summer BOUM cruise, Biogeosciences, 8, 2159¿2177, doi:10.5194/bg-8-2159-2011.
Raybaud, V., Nival, P., Mousseau, L., Gubanova, A., Altukhov, D., Khvorov, S., Ibanez, F., Andersen, V., 2008. Short term changes in zooplankton community during the summer-autumn transition in the open NW Mediterranean Sea: species composition, abundance and diversity. Biogeosciences 5, 1765¿1782. https://doi.org/10.5194/bg-5-1765-2008.
Ribera d'alcala, M., Conversano, F., Corato, F., Licandro, P., Mangoni, O., Marino, D., Mazzocchi, M.G., Modigh, M., Montresor, M., Nardella, M., Saggiomo, V., Sarno, D., Zingone, A., 2004. Seasonal patterns in plankton communities in a pluriannual time series at a inshore Mediterranean site (Gulf of Naples): an attempt to discern recurrences and trends. Scientia Marina 68, 65-83.
Vernette C, Henry N, Lecubin J, de Vargas C, Hingamp P, and Lescot M The Ocean Barcode Atlas: a web service to explore the biodiversity and biogeography of marine organisms. (2021), Mol Ecol Resour. 21: 1347-1358. https://doi.org/10.1111/1755-0998.13322
Chapitre de livre:
Coppola L., Boutin J., Gattuso J.-P., Lefèvre D. & Metzl N. (2020). The carbonate system in the Ligurian Sea. In: Migon C., Nival P. & Sciandra A. (Eds.), The Mediterranean Sea in the era of global change (volume 1) - Evidence from 30 years of multidisciplinary study of the Ligurian sea, pp. 79-104. London: ISTE Science Publishing LTD. https://doi.org/10.1002/9781119706960.ch4
Thèses:
Feliu Guillermo (2021). Study of the mesozooplankton community structure and trophic role in the Mediterranean Sea during two large scale oceanographic surveys PEACETIME and MOOSE in 2017. PhD Thesis, Université Aix-Marseille
Maili K.: Étude de la variabilité océanique du bassin algérien. Thèse Sorbonne Universités. Encadrants : L. Mortier et P. Testor (prévue 2022)
Margirier F. : Etude de la variabilité physique et biogéochimique des masses d'eaux en Mer Méditerranée nord occidentale. Thèse Sorbonne Universités. Encadrants : L. Mortier et P. Testor (2018)

 , BOSSE Anthony
, BOSSE Anthony 