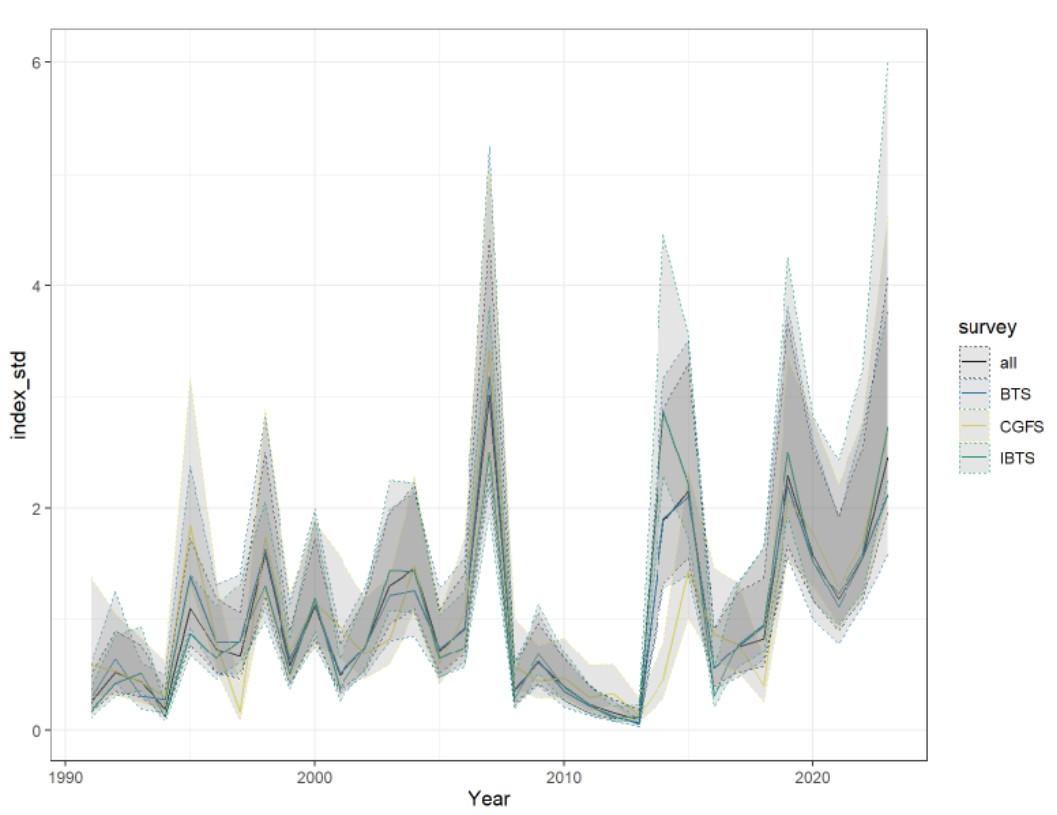
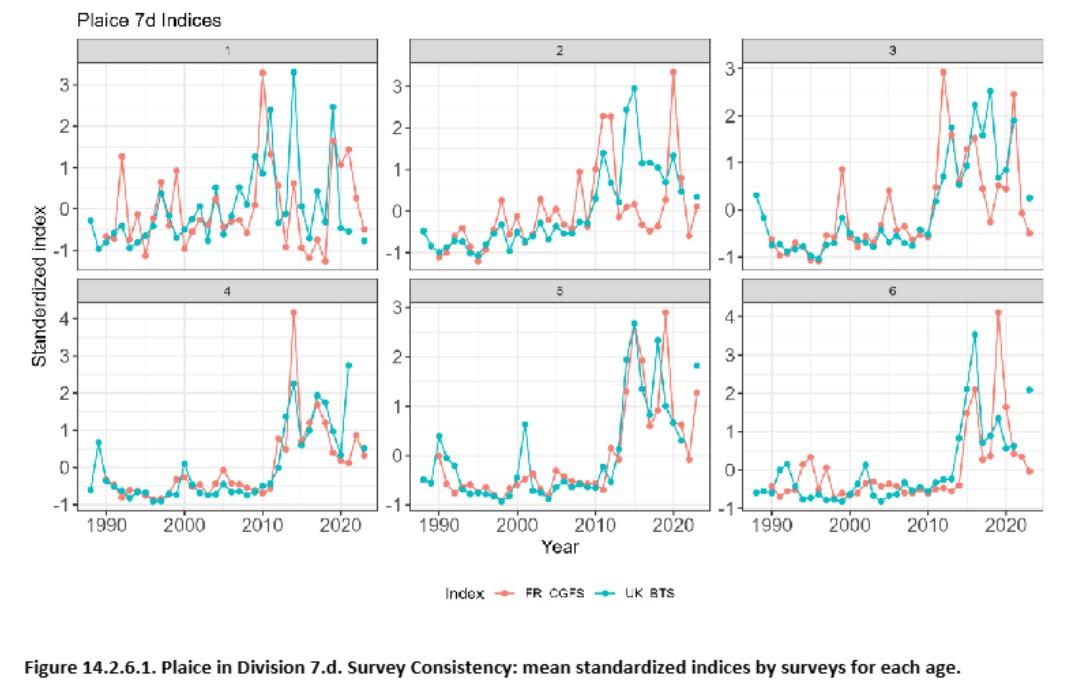
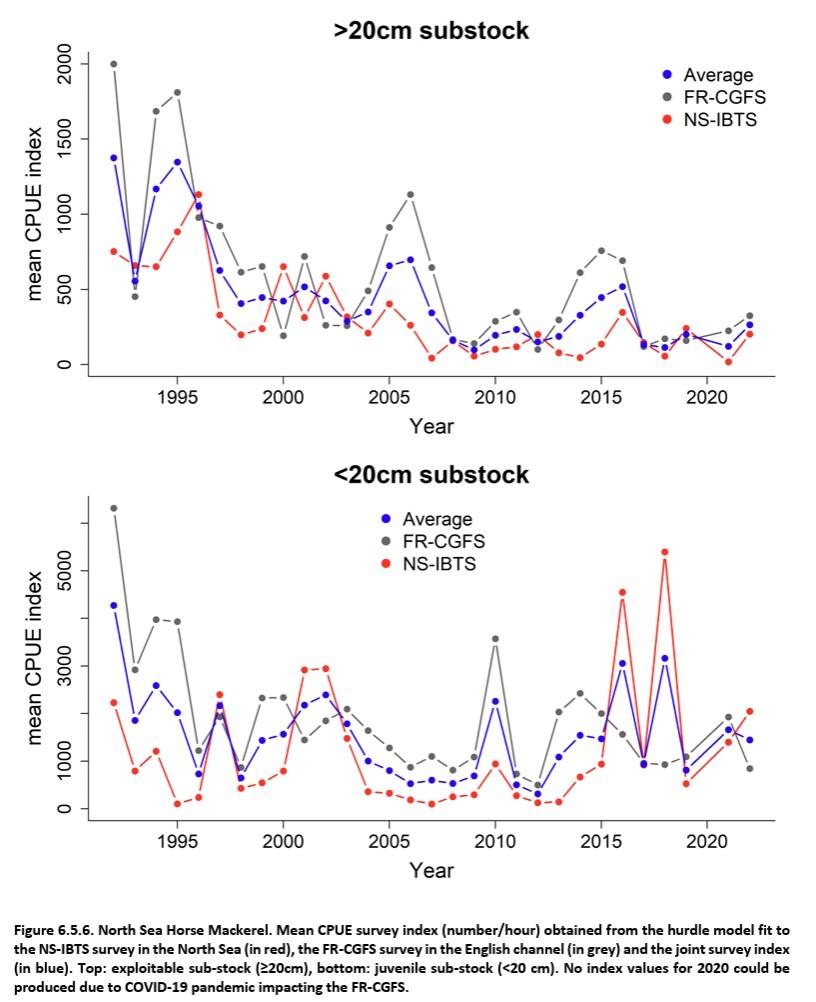
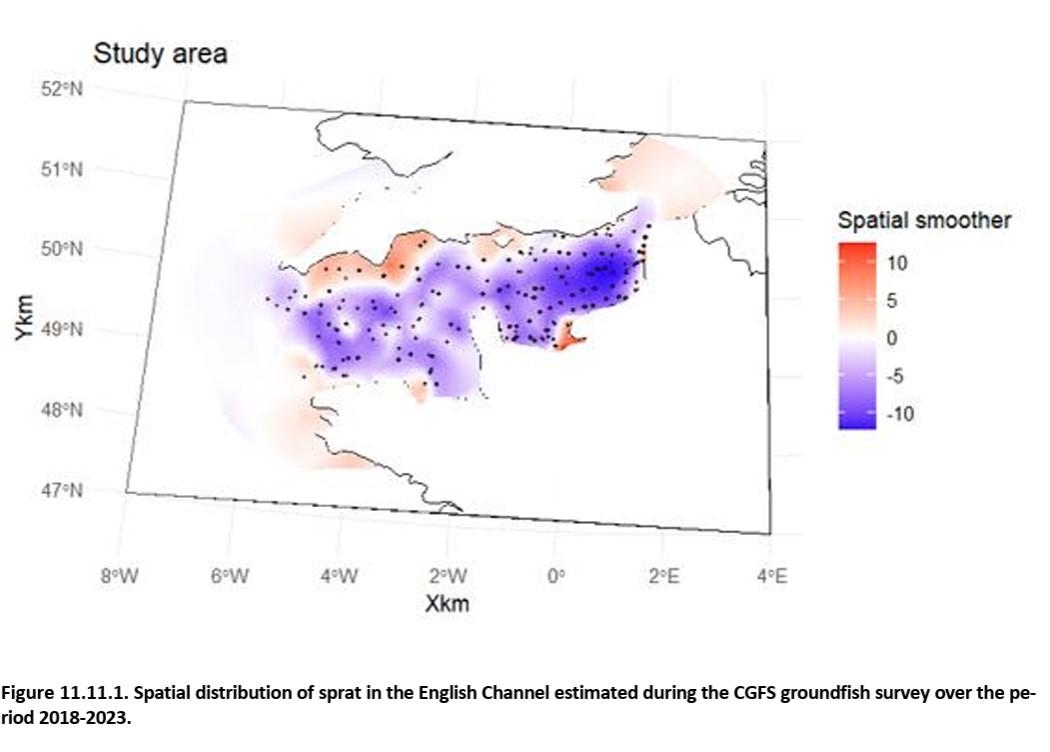
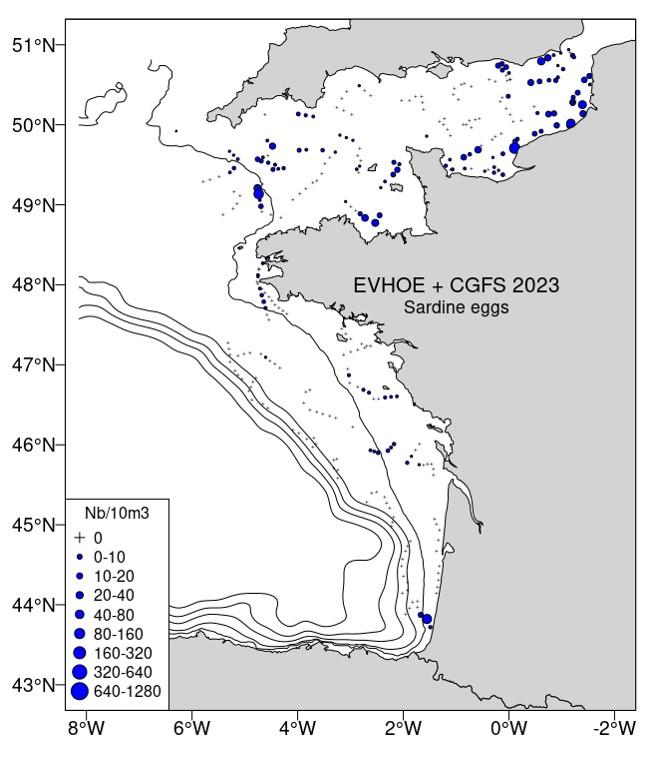
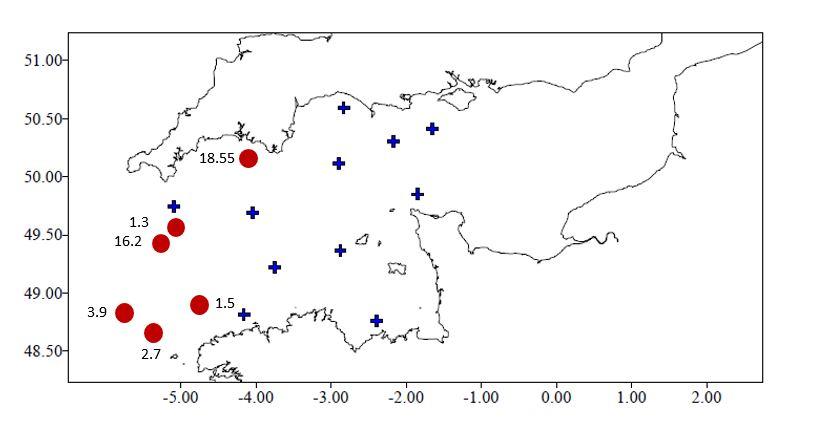
Références des rapports techniques
Doray Mathieu
,
Daures Fabienne,
Ernande Bruno,
Girardin Raphael,
Giraldo Carolina,
Halouani Ghassen,
Huret Martin,
Lacroix Denis,
Lapègue Sylvie, Lefebvre Sebastien,
Pernet Fabrice,
Marchal Paul,
Travers-Trolet Morgane (2025).
Rapport du projet FORESEA 2050 : scénarios de contribution des produits de la mer à la sécurité alimentaire française en 2050. RBE/HALGO/EMH 25-03.
https://doi.org/10.13155/105669 Le Corre Pauline (2024). Distribution modelling of benthic invertebrate biodiversity of the western European continental shelves and impact of climate changes. 5th-year Internship Report. INSA Lyon.
Jadaud Angelique (2024).
Note interne. Responsables des campagnes de suivi halieutique. CAMPAGNES_Responsables_V10_2024_06_28.docx.
Cornou Anne Sophie (2023).
Guide d'utilisation de l'application Vivaldi2.0. Exemple format datras avec données CGFS.
Huguet Antoine, Marais Lucas (2023).
Exigences fonctionnelles et technique / Proposition d'Architecture Technique pour les livraisons de données aux Conventions des Mers Régionales. SIMM - Système d'Information Milieu Marin. Cahier des Charges. ODE/VIGIES 23-17.
https://archimer.ifremer.fr/doc/00876/98786/ Blanchard Ariane, Laran Sophie, Williams Gary, Van Canneyt Olivier, Genu Mathieu, Sanchez Thierry, Doremus Ghislain, Gamelin Pierre-Louis (2023).
Synthèse des données de l’Observatoire PELAGIS au sein du PNM des estuaires picards et de la mer d’Opale. Rapport pour le PNM EPMO (OFB-22-0565). 61 pp.
https://archimer.ifremer.fr/doc/00848/95984/ICES (2023).
Stock Annex: Striped red mullet (Mullus surmuletus) in Subarea 4 and divisions 7.d and 3.a (North Sea, eastern English Channel, Skagerrak and Kattegat). ICES Stock Annexes. 29 pp.
https://doi.org/10.17895/ices.pub.23578803Duc Emilie,
Huguet Antoine (2023).
Rapport final - Bilan du chantier « Collecte des données DCSMM-CMR-DSF ». Evaluation du cycle 3 de la Directive Cadre « Stratégie pour le Milieu Marin » - Convention des Mers Régionales – Documents Stratégiques de Façades. ODE/VIGIES/23-06.
https://archimer.ifremer.fr/doc/00834/94574/Régimbart Amélie (2023).
Bilan de l’optimisation des campagnes halieutiques réalisée dans le cadre du programme de surveillance de la directive cadre stratégie pour le milieu marin (DCSMM) - année 2022. Rapport scientifique, Ifremer ODE/VIGIES/DCSMM, 40p.
https://archimer.ifremer.fr/doc/00834/94623/ Dubost Gaspard, Elliott Sophie, Rivot Etienne, Acou Anthony, Beaulaton Laurent,
Deleys Noémie, Reveillac Elodie (2023).
Rapport du projet « MigrenMer » : Synthèse et valorisation des connaissances disponibles sur les migrateurs amphihalins en mer. Rapport scientifique du pole MIAME. 81 p. + annexes.
https://archimer.ifremer.fr/doc/00878/99026/Régimbart Amélie (2023).
Bilan de l’optimisation des campagnes halieutiques réalisée dans le cadre du programme de surveillance de la directive cadre stratégie pour le milieu marin (DCSMM) - année 2022. Rapport scientifique, Ifremer ODE/VIGIES/DCSMM, 40p.
https://archimer.ifremer.fr/doc/00834/94623/ Le Gac Romane (2023). Participation à l’évaluation des impacts anthropiques sur les communautés benthiques côtières au LEBCO. Rapport d’alternance de licence professionnelle Technicien Supérieur Sciences Halieutiques. Université du Littoral Côte d'Opale.
Régimbart Amélie (2022).
Bilan de l’optimisation des campagnes halieutiques réalisée dans le cadre du programme de surveillance de la directive cadre stratégie pour le milieu marin (DCSMM) - année 2021. Rapport scientifique, Ifremer ODE/VIGIES/DCSMM, 40p.
https://archimer.ifremer.fr/doc/00783/89515/ Simonet Lea (2022). Trends in demersal populations' growth and condition: Bay of Biscay and Celtic Sea case studies.
Liefmann Stephanie, Dauvin Jean-Claude, Delahaye Marc, Domingo Camille, Evrard Manuel, Guyet-Grenet Valérie, Manner Arnauld, Mouillard Romain, Prévalet Solène, Robin Jean-Paul,
Vigneau Joel (2022).
Effets des engins de pêche sur les fonds marins de Manche-Est et du golfe normand-breton. Rapport final du projet IPREM - Impact des engins de Pêche sur les fonds marins et la Résilience Ecologique du Milieu (2021-2022).
https://doi.org/10.13155/89603Vincent D., Goberville E., Vilain M., Goffart A. (2022).
Campagnes halieutiques optimisées pour la DCSMM. Analyse du dispositif de surveillance pour répondre aux besoins du programme thématique « Habitats pélagiques ».
https://archimer.ifremer.fr/doc/00848/95995/Régimbart Amélie (2022).
Bilan de l’optimisation des campagnes halieutiques réalisée dans le cadre du programme de surveillance de la directive cadre stratégie pour le milieu marin (DCSMM) - année 2021. Rapport scientifique, Ifremer ODE/VIGIES/DCSMM, 40p.
https://archimer.ifremer.fr/doc/00783/89515/ Moulin Nina (2021). Distribution et exploitation du maquereau, du chinchard et du sprat par les pêcheries françaises dans l’Atlantique Nord-Est. Mémoire de Master 1, Sciences de la mer et du littoral, Mention Biologie, Sciences Halieutiques et Aquacoles. Université de Bretagne Occidentale.
Coulon Noémie (2021). Rareté fonctionnelle des communautés de poissons de la côte nord-est de l'Atlantique / Functional rarity of the Northeast Atlantic coastal fish communities. Master’s Thesis. Year 2020-2021, Master's Degree in Marine Sciences.
Baudrier Jerome (2020).
Bilan de l’optimisation des campagnes halieutiques réalisée dans le cadre du programme de surveillance de la directive cadre stratégie pour le milieu marin (DCSMM) - année 2019. Rapport scientifique, Ifremer ODE/VIGIES/DCSMM, 48 p.
https://archimer.ifremer.fr/doc/00609/72095/ Jouandet Marie-Pâule, Lebourg Emeline, Bruault Marie, Dedecker Claire, Louchart Arnaud, Gomez Fernando, Nowaczyck Antoine, Wacquet Guillaume, Lizon Fabrice, Crouvoisier Muriel, Cornille Vincent, Lefebvre Kévin, Bouchaud Fanny, Lécuyer Eric, Mériaux Xavier, Cauvin Arnaud, Didry Morgane, Delabre Jessica, Duflos Marie, Artigas Luis Felipe (2020). Résultats des campagnes océanographiques 2017- 2019 en Mers Celtiques et Manche Mer du Nord. Test pour la mise en place du Programme de Surveillance DCSMM. [PHYCO 2017 - PELRAD 2017 - ECOPEL 2018 leg 1 et leg 2 - CGFS 2018 - IBTS 2018 et 2019].
Baudrier Jerome (2020).
Programmes de surveillance DCSMM 1er cycle : « Espèces commerciales » et « Poissons-céphalopodes (SP3-SP5) » : synthèse des résultats et priorisation des dispositifs proposés pour la surveillance second cycle. ODE/VIGIES/DCSMM, 43 p.
https://archimer.ifremer.fr/doc/00601/71294/ Rivet Jehanne (2020).
Régionalisation de l'espace maritime pour sa planification et gestion: le cas de la baie de Seine. Rapport de stage de 2ème année de Master. Mention Biodiversité, Ecologie, Evolution (BEE). Parcours FOnctionnement et Gestion des Ecosystèmes Marins (FOGEM). Université de Lille.
https://archimer.ifremer.fr/doc/00704/81578/Fontaine Virginie (2020). Dynamique et structure du réseau trophique planctonique en Manche-Est. Rapport de Stage de 2éme année de Master Biodiversité, Ecologie, Evolution (BEE). Parcours FOnctionnement et Gestion des Ecosystèmes Marins (FOGEM). Université de Lille.
Fontaine Virginie (2020). Dynamique et structure du réseau trophique planctonique en Manche-Est. Rapport de Stage de 2éme année de Master Biodiversité, Ecologie, Evolution (BEE). Parcours FOnctionnement et Gestion des Ecosystèmes Marins (FOGEM). Université de Lille.
Jouandet Marie-Paule, Petit Laëtitia, Duflos Marie,
Wacquet Guillaume, Dedecker Claire, Aubert Anaïs, Rombouts Isabelle, Mialet Benoit, Lebourg Emeline, Bruault Marie, Gomez Fernando, Nowaszyk Antoine, Delarbre Jessica, Cornille Vincent, Lecuyer Eric, Didry Morgane, Lizon Fabrice, Artigas Luis Felipe (2019).
Mise en oeuvre du programme de surveillance "Habitats Pélagiques" : test de mise en place des campagnes saisonnières et calcul d’indicateurs sur le site atelier Manche Mer du Nord et Mers Celtiques. Rapport sur les campagnes DCSMM 2017-2019 – SRM MMN . 127p.
Gravier Nathan (2019).
Dynamique spatiale et interannuelle des communautés mésozooplanctoniques en Manche Orientale en lien avec l’introduction récente d’une espèce non-indigène : Mnemiopsis leidyi.
https://archimer.ifremer.fr/doc/00504/61551/Auber Arnaud,
Laffargue Pascal (2019).
Compte rendu de participation au Groupe d'experts du CIEM consacré aux campagnes scientifiques (WGIBTS). Den Helder (Pays-Bas), du 01 au 05 avril 2019. RBE/ederu/2019/CR14.
Regimbart Amelie, Guitton Jérôme, Le Pape Olivier (2018).
Zones fonctionnelles pour les ressources halieutiques dans les eaux sous souveraineté française. Deuxième partie : Inventaire. Rapport d'étude. Les publications du Pôle halieutique AGROCAMPUS OUEST n°46, 175 p.
https://archimer.ifremer.fr/doc/00652/76437/ Drogou Mickael, Laurec Alain, Bissery Claire,
Mahevas Stephanie,
Demaneche Sebastien, Begot Eric, Laurens Martial,
Le Goff Ronan, de Pontual Helene
,
Talidec Catherine, Weiss Jerome
, Allanic Guillaume,
Barone Herve, Bazet Mathilde,
Bouche Ludovic,
Le Bourdonnec Pierre,
Coupeau Yann,
Dimeet Joel,
Duhamel Erwan,
Fauvel Christian, Le Gall Marie-Annik,
Garren Francois,
Latrouite Armelle,
Lespagnol Patrick,
Marc Emilie,
Martin Stephane,
Le Mestre Sophie,
Nedelec Daniel,
Pennors Laurence,
Le Roy Didier,
Le Ru Loic,
Salaun Michele,
Villanueva Ching-Maria (2017).
Projet Bargip. Action Données professionnelles. Rapport final mars 2017. RST-RBE/STH/LBH/17-002.
Le Moigne Morgan,
Lamoureux Alice,
Brun Melanie (2017).
Répondre à l’estimation des effets de la mesure d’interdiction des sacs plastiques à usage unique en caisse. juillet 2016. Les données de la surveillance à disposition dans la base de données Quadrige². ODE/VIGIES/ MLM.
Baudrier Jerome (2017).
Bilan de l’optimisation des campagnes halieutiques - Programme de surveillance DCSMM, année 2016. ODE/VIGIES/DCSMM/17-009.
Aubert Anaïs, Thibault Delphine (2017).
Guide DCSMM d’aide à la détermination des principaux types/espèces de macro- et mégazooplancton gélatineux. Programme de Surveillance DCSMM. 70p.
https://doi.org/10.13155/50398Mc Lean Matthew James,
Auber Arnaud, Mouillot David (2017).
Projet RESTICLIM. Compte-rendu du 1er comité de thèse "Réponses fonctionnelles des communautés de poissons aux gradients environnementaux en Baie de Somme, Manche orientale et Mer du Nord." /
RESTICLIM project. Report of the 1st committee of thesis. Délivrable n°2.
Auber Arnaud (2016).
Effets de la PêChe et du CLImat sur la biodiversité de PoiSsons en Manche et MEr du Nord. Rapport intermédiaire - projet ECLIPSE.
Robert Marianne,
Foucher Eric,
Drogou Mickael (2016).
Compte rendu de participation au groupe d’experts du CIEM pour l’évaluation des stocks des mers celtiques (WGCSE). Copenhague, 3-13 mai 2016. RBE/EDERU/2016/CRGT10.
ICES (2016). Report of the Working Group on Elasmobranch Fishes (WGEF). 15-24 June 2016. Lisbon, Portugal. ICES CM/ACOM:20.
Auber Arnaud (2016).
Rapport intermédiaire – projet ECLIPSE. Effets de la PêChe et du CLImat sur la biodiversité de PoiSsons en Manche et MEr du Nord.
Auber Arnaud (2016).
Effets de la PêChe et du CLImat sur la biodiversité de PoiSsons en Manche et MEr du Nord. Rapport intermédiaire - projet ECLIPSE.
Stephan E, Gadenne Helene, Meheust E, Jung Jl (2015). Projet RECOAM. Etude de cinq espèces de raies présentes dans les eaux côtières d'Atlantique et de Manche. Rapport final.
Saunier Jean-Baptiste, Leprieur Fabrice (2015).
Constat radiologique « Nord – Normandie ». Rapport méthodologique relatif aux compartiments aquatiques continental et marin. PRP-ENV/SESURE/2015-01.
https://archimer.ifremer.fr/doc/00640/75217/Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Morin Jocelyne,
Brind'Amour Anik, Fisson Cédric, Bacq Nicolas, Boulet M, Guillerme D, Berthele Olivier,
Martin Jocelyne (2015).
Peuplements halieutiques en estuaire de Seine et baie de Seine orientale - Distribution et évolution des abondances des principales espèces capturées au chalut à perche lors des campagnes scientifiques réalisées par l’Ifremer entre 1995 et 2010.
Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Foucher Eric, Blanchard Michel,
Cugier Philippe,
Desroy Nicolas,
Dreanno Catherine, Fauchot Juliette,
Fifas Spyros,
Guyader Olivier, Jean Frederic, Le Gallic Bertrand,
Riou Philippe,
Schapira Mathilde, Thiebaut Eric (2015).
Le projet ANR-COMANCHE 2010-STRA-010. Interactions écosystémiques et impacts anthropiques dans les populations de COquilles Saint-Jacques (Pecten maximus) de la MANCHE. Rapport scientifique final.
https://archimer.ifremer.fr/doc/00251/36236/ Lorance Pascal,
Berthou Patrick,
Bertrand Jacques,
Dintheer Christian,
Ernande Bruno,
Foucher Eric,
Huret Martin,
Travers-Trolet Morgane,
Trenkel Verena (2014).
Rapport du groupe de travail Campagnes halieutiques en Manche. Groupe de travail interne RBE, réflexion sur les campagnes halieutiques en Manche, février-mars 2014.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Villanueva Ching-Maria,
Ernande Bruno, Baheux Mickael, Lefevbre Sébastien (2014).
Écologie fonctionnelle et relations trophiques de poissons: Protocoles de collecte et analyse des échantillons en Manche Orientale. R.INT.STH.HMMN/RBE/LBH.RH LAB RESEAUX TROPHIQUES 2014-1.
Marchal Paul, Bartelings Heleen, Bastardie François, Batsleer Jurgen, Delaney Alyne,
Girardin Raphael, Gloaguen Pierre, Hamon Katell, Hoefnagel Ellen, Jouanneau Charlène,
Mahevas Stephanie, Nielsen Rasmus, Piwowarczyk Joanna, Poos Jan-Jaap, Schulze Torsten, Rivot Etienne, Simons Sarah, Tidd Alex,
Vermard Youen,
Woillez Mathieu (2014).
Mechanisms of change in human behaviour. WP2.3 D.2.3.1.
https://archimer.ifremer.fr/doc/00223/33377/ Coppin Franck,
Lazard Coline (2013).
Fiche Roscop : CGFS 2013. Compte-rendu de Campagne à la Mer sur un navire hauturier et côtier de la TGIR Flotte océanographique française.
Coppin Franck,
Le Roy Didier (2013).
CGFS 2014. Campagne expérimentale de chalutage en Manche orientale. Channel Ground Fish Survey. Programme scientifique.
Vacherot Jean-Philippe (2012).
Rapport d'essais de l'utilisation de VECOC lors de la campagne CGFS 2012. N/O Gwen-Drez du 26 au 31 octobre 2012. RBE/STH/LTBH 2012-07.
Curd Amelia (2011). Conférence mondiale sur la biodiversité (26-30/09/11) - Aberdeen, Scotland (U.K.).
Rocklin Delphine, Pelletier Dominique, Levrel Harold (2011). Projet SINP (volet mer) - Test des indicateurs institutionnels de biodiversité marine et côtière.
Trenkel Verena (2011). Rapport de mission aux groupes CIEM WGISUR et WKCATDAT / Working Group on Integrating Surveys for the Ecosystem Approach (WGISUR) et Workshop on Cataloguing Data requirements from surveys for the EAFM (WKCATDAT).
Trenkel Verena, Leonardi Sophie, Vignot Celine (2011). Intégration d?indicateurs concernant les ressources halieutiques et les activités de pêche dans le tableau de bord des mers françaises.
Vermard Youen (2011). Compte rendu de participation au Groupe Mer du Nord et Skagerrak (WGNSSK)
Fossat J,
Pelletier Dominique, Levrel Harold (2010).
Projet Système d'Information sur la Nature et les Paysages, volet mer (SINP-mer). Synthèse et typologie des indicateurs institutionnels de biodiversité marine et côtière. NA : A110204.
Biseau Alain (2010). Situation en 2010 des ressources exploitées par les flottilles françaises.
Mahe Jean-Claude, Bellail Robert, Drogou Mickael, Mahe Kelig (2010). Compte rendu de participation au groupe de travail "Nouvelles espèces" (CIEM/WGNEW).
Vermard Youen (2010). Compte rendu de participation au Groupe Mer du Nord et Skagerrak (WGNSSK).
Bertrand Jacques, Brind'Amour Anik, Cochard Marie-Laure, Coppin Franck, Leaute Jean-Pierre, Lorance Pascal, Mahe Jean-Claude, Morin Jocelyne, Poulard Jean-Charles, Rochet Marie-Joelle, Schlaich Ivan, Souplet Arnauld, Trenkel Verena, Verin Yves (2009). Grands invertébrés et poissons observés par les campagnes scientifiques. Bilan 2007.
Fossat Julia, Pelletier Dominique, Levrel Harold (2009). Projet SINP - Synthèse des indicateurs institutionnels de biodiversité marine et côtière.
Morin Jocelyne, Bertrand Jacques, Cochard Marie-Laure, Coppin Franck, Leaute Jean-Pierre, Lobry Jeremy, Mahe Jean-Claude, Poulard Jean-Charles, Rochet Marie-Joelle, Schlaich Ivan, Souplet Arnauld, Trenkel Verena, Vaz Sandrine, Verin Yves (2009). L'état des communautés exploitées au large des côtes de France. Application d'indicateurs à l'évaluation de l'impact de la pêche. Bilan 2004 - Edition 2009.
Trenkel Verena, Berthele Olivier, Lorance Pascal, Bertrand Jacques, Brind'Amour Anik, Cochard Marie-Laure, Coppin Franck, Leaute Jean-Pierre, Mahe Jean-Claude, Morin Jocelyne, Rochet Marie-Joelle, Salaun Michele, Souplet Arnauld, Verin Yves (2009). Grands invertébrés et poissons observés par les campagnes scientifiques. Atlas de distribution. Bilan 2008.
Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Carpentier Andre,
Coppin Franck, Curet Lucie, Dauvin Jean-Claude, Delavenne Juliette, Dewarumez Jean-Marie, Dupuis Ludovic,
Foveau Aurélie, Garcia Clément, Gardel Laure, Harrop Stuart, Just Roger, Koubbi Philippe, Lauria Valentina, Martin Corinne, Meaden Geoff,
Morin Jocelyne, Ota Yoshi,
Rostiaux Emilie, Smith Bob, Spilmont Nicolas,
Vaz Sandrine,
Villanueva Ching-Maria,
Verin Yves, Walton Joanne, Warembourg Caroline (2009).
Atlas des Habitats des Ressources Marines de la Manche Orientale - CHARM II /
Channel Habitat Atlas for marine Resource Management - CHARM II. PROGRAMME INTERREG 3A.
https://archimer.ifremer.fr/doc/00000/7377/ Huet Jerome, Morizur Yvon, Mahe Kelig (2008). Analyse d?estimation de l?âge individuel du bar (Dicentrarchus labrax) du stock Manche Est (Division CIEM VIId), stock Manche Ouest - mer Celtique (Division CIEM VIIe, h) et du stock golfe de Gascogne (Division CIEM VIIIa, b).
Lizaud Oceann, Mahe Kelig (2008). Protocole technique d?estimation de l?âge individuel de la cardine (Lepidorhombus whiffiagonis) de mer Celtique et du golfe de Gascogne (Divisions VII g, h, j et VIII a,b du CIEM).
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Prouzet Patrick (2008). Rapport de restitution du Programme : Démarche intégrée pour une gestion écosystémique des ressources halieutiques - DEMOSTEM.
Bertrand Jacques,
Cochard Marie-Laure,
Coppin Franck,
Leaute Jean-Pierre, Lobry Jeremy,
Mahe Jean-Claude,
Morin Jocelyne, Poulard Jean-Charles, Rochet Marie-Joelle,
Schlaich Ivan, Souplet Arnauld,
Trenkel Verena,
Verin Yves (2007).
Poissons et invertébrés au large des côtes de France. Indicateurs issus des pêches scientifiques. Bilan 2004. IFREMER, NANTES, EMH : 07-001.
https://archimer.ifremer.fr/doc/00000/2327/ Delpech Jean-Paul, Mahe Kelig, Mehault Sonia, Rostiaux Emilie, Biais Gerard, Coppin Franck, Vaz Sandrine (2007). Évaluation des ressources halieutiques par les campagnes scientifiques françaises - Façade "Manche est" et façade "Loire-Gironde".
Coppin Franck, Le Roy Didier, Schlaich Ivan (2006). Guide du chef de mission de la campagne CGFS (Channel Ground Fish Survey).
Dufour Jean-Louis, Mahe Kelig (2006). Protocole technique d?estimation de l?âge individuel de la morue (Gadus morhua) de mer du Nord et Manche orientale (Divisions CIEM IVb, c et VIId).
Dufour Jean-Louis, Mahe Kelig (2006). Protocole technique d?estimation de l?âge individuel du merlan (Merlangius merlangus) en mer du Nord et Manche Est (Divisions CIEM IVb, IVc, VIId).
Dufour Jean-Louis, Mahe Kelig (2006). Analyse de la méthode d?estimation de l?âge individuel du merlan (Merlangius merlangus) en mer du Nord et Manche Est (Divisions CIEM IVb, IVc, VIId).
Leroy Didier, Mahe Kelig (2006). Analyse d?estimation de l?âge individuel du rouget barbet de roche (Mullus surmuletus) en Manche orientale et sud mer du Nord (divisions CIEM VIId et IVc).
Leroy Didier, Mahe Kelig (2006). Protocole technique d?estimation de l?âge individuel du rouget barbet de roche (Mullus surmuletus) en Manche orientale et sud mer du Nord (divisions CIEM VIId et IVc).
Manten Marie-Line, Mahe Kelig (2006). Protocole technique d?estimation de l?âge individuel de la plie (Pleuronectes platessa) en mer du Nord et Manche Est (Divisions CIEM IVb, IVc, VIId).
Manten Marie-Line, Mahe Kelig (2006). Protocole technique d?estimation de l?âge individuel de la sole de mer du Nord et manche Est (Divisions CIEM IVb,c et VIId).
Bertrand Jacques, Cochard Marie-Laure, Coppin Franck, Le Pape Olivier, Mahe Jean-Claude, Morin Jocelyne, Poulard Jean-Charles, Rochet Marie-Joelle, Schlaich Ivan, Souplet Arnauld, Trenkel Verena, Verin Yves (2005). Poissons et invertébrés au large des côtes de France : Indicateurs issus des pêches scientifiques. Bilan 2002.
Mahe Kelig, Destombe A., Coppin Franck, Koubbi P., Vaz Sandrine, Le Roy Didier, Carpentier Audrey (2005). Le rouget barbet de roche Mullus surmuletus (L. 1758) en Manche orientale et mer du Nord.
Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Carpentier Andre,
Coppin Franck, Dauvin Jean-Claude,
Desroy Nicolas, Dewarumez Jean-Marie, Eastwood Paul D.,
Ernande Bruno, Harrop Stuart, Kemp Zarine, Koubbi Philippe, Williams Nigel Leader,
Lefebvre Alain, Lemoine Michel,
Loots Christophe, S. Martin Corinne, Meaden Geoff J., Ryan Nick, Tan Lei,
Vaz Sandrine, Walkey Mike (2005).
CHARM : Atlas des Habitats des Ressources Marines de la Manche Orientale. PROJET CHARM - RAPPORT FINAL (ATLAS ET RAPPORT TECHNIQUE).
https://archimer.ifremer.fr/doc/00000/2350/ Forest Andre (2001).
Ressources halieutiques hors quotas du Nord Est Atlantique : bilan des connaissances et analyse de scénarios d'évolution de la gestion. Contrat Ifremer/MAPA – Réf. 99-I1-03-01.
https://archimer.ifremer.fr/doc/00000/1201/Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Woillez Mathieu (2001).
Utilisation de la stabilité temporelle de la répartition spatiale de certaines ressources de la mer du Nord pour leur cartographie.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Coppin Franck,
Delpech Jean-Paul,
Dufour Jean-Louis (2001).
Campagnes d'évaluation des Ressources Halieutiques en mer du Nord et Manche Orientale. Volume I Présentation des campagnes I.B.T.S. C.G.F.S. Volume II. Résultats des campagnes I.B.T.S. 99, I.B.T.S. 2000 C.G.F.S. 99, C.G.F.S. 2000. Contrat BIOECO n° 98/058.
Verin Yves,
Carpentier Andre (1997).
Campagnes expérimentales de chalutage en mer du nord et manche orientale. Volime. I : les campagnes IBTS 1995 & 1996. Volume II : les campagnes CGFS 1995 & 1996.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Verin Yves,
Carpentier Andre (1996).
Campagnes expérimentales de chalutage en Mer du Nord et Manche Orientale. Rapport préliminaire. Volume I Les campagnes IBTS. Volume II Les campagnes CGFS.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Guillot Sylvia (1996). Traitement informatique de données de campagnes à la mer pour analyse statistique des peuplements halieutiques de la Baie de Seine. Rapport de Dess Informatique Appliqué aux Sciences de la Vie. Université Paris V.
Delcour Sophie (1996). Répartition et abondance du Grondin rouge (Aspitrigla cuculus) en Manche Orientale. Campagne CGFS 95. Traitement informatique & Analyse des résultats.
Ifremer (1995). Réunion du département " Ressources Halieutiques ". Nantes - 5 et 6 décembre 1995.
Bennet D, Casey J, Dare P, Dawson W, Flatman S, Hulme T, Macer C, Millner R, Pickett G, Vince M, Lorance Pascal, Souplet A, Giret M, Morin Jocelyne, Berthou A, Latrouite Daniel, Morizur Yvon, Bossy S, Ozanne S (1993). Identification biogéographique des principaux stocks exploités en Manche, relations avec ceux des régions voisines.

 , LE ROY Didier , MARTIN-BAILLET Victor
, LE ROY Didier , MARTIN-BAILLET Victor