Stock assessment
The CGFS contributes for the Eastern Channel, to the production of abundance indices for red mullet (Mullus surmuletus, Fig 1), plaice (Pleuronectes platessa, Fig 2), or cephalopods (Sepia officinalis, Fig 3); collecting information on the stage of maturity of the main commercial species (whiting (Merlangius merlangus), cod (Gadus morhua)) in order to know the proportion of mature individuals at each age. This type of information is essential for calculating the quantity of fish of reproductive age (spawning stock biomass), the level of which is one of the main indicators of the stock's health.
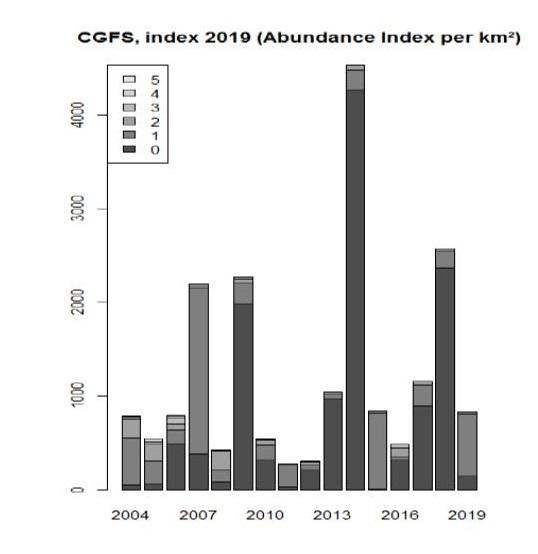
Figure 1: Extract from the last report of the WGNSSK group (https://www.ices.dk/community/groups/Pages/WGNSSK.aspx) and which summarizes the main results of models for the assessment of red mullet in the Channel.
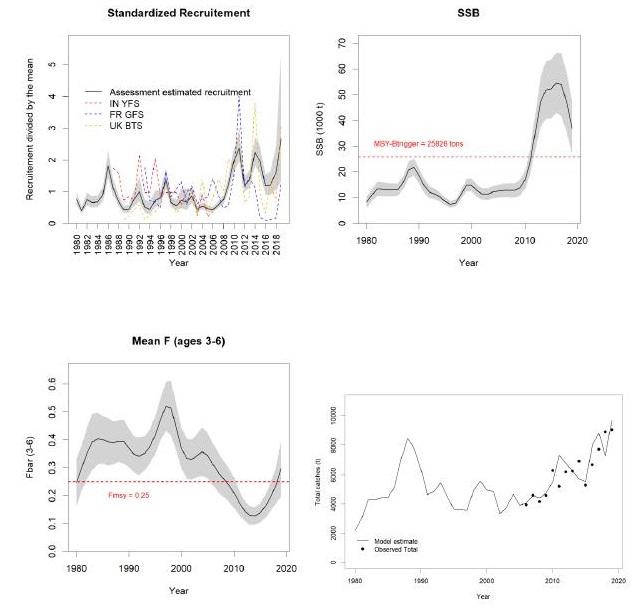
Figure 2: Extract from the latest WGNSSK 2020 report (https://www.ices.dk/community/groups/Pages/WGNSSK.aspx) and which summarizes the main results of assessment models on Channel plaice.
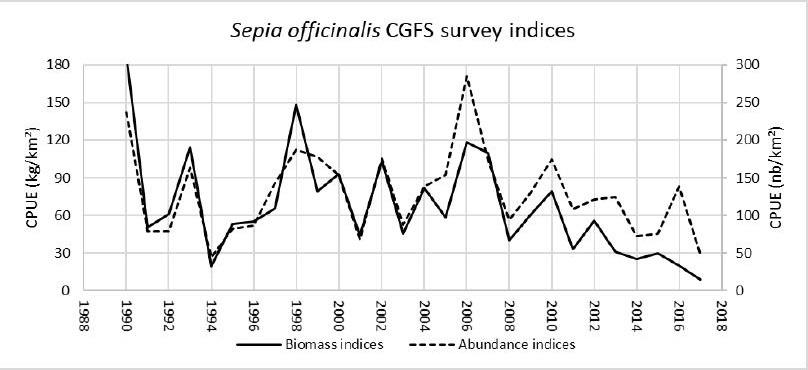
Figure 3 : Extract from the latest report of the Working Group on Cephalopod Fisheries and Life History (WGCORP 2020) https://www.ices.dk/community/groups/Pages/WGCEPH.aspx
The abundance indices calculated from scientific surveys can be used in models such as SURBAR to estimate fluctuations in biomass, mortality, and recruitment. This type of analysis is based only on survey data and for the first time in 2020, the WGNSSK working group tested its application on the CGFS data of mullet in order to be able to compare the outputs of different stock assessment models. (Fig 4). In addition, the data collected during the survey makes it possible to know the demographic size-structures for all the species, the inter-annual fluctuations in population indicators such as abundance, biomass, average size and average weight. The CGFS also provides elements to study possible spatio-temporal modifications of the ecosystem.
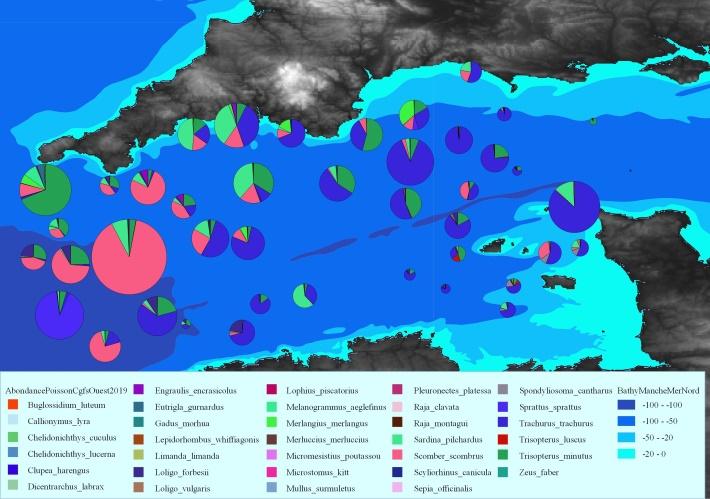
Figure 4: Main species composition of the catch in the Western English Channel during CGFS 2019
Ecosystem approach
Sampling the entire ecosystem makes it possible to respond to the demands of the Marine Strategy Framework Directive (MSFD, Figure 5), both for the search for a definition of good ecological status, but also for monitoring of the various descriptors characterizing it, and in particular the descriptors:
1: Biological diversity, habitat quality, distribution and abundance of species
2: Level of presence of non-native species
5: Human-induced eutrophication
4: Food web, diversity and abundance of its elements
6: Seabed integrity, structure and function of benthic ecosystems

Figure 5: MSFD sampling stations
Food-web structure & functioning
Depth gradient on the resource use of a fish community from a semi- enclosed sea: Depth is one of the environmental variables influencing the structure of marine food webs by directly or indirectly influencing benthic-pelagic coupling and predator-prey relationships. In shallow waters, the high degree of connectivity between pelagic and benthic networks results in complex systems with multiple inter- actions. Digestive tract (DT) and stable isotope (SI) analyses were used to investigate depth-related changes in feeding patterns for 33 fish species (eastern English Channel [EEC]) collected between 5 m and 80 m depth. Fish species were first arranged into functional groups based upon trophic and habitat similarities. DTs were used to determine the general topology of the food web and SIs were used to estimate the contributions of different sources to fish diets. At the scale of the aggregated EEC food web, the main food sources for all groups were of benthic origin (> 50%). The aggregated food web was then used as a template to explore the influence of depth on resource use by predatory fish. Mixing models including depth as a continuous covari- ate successfully untangled and identified different feeding strategies among functional groups. In shallow waters, fish species benefited from both pelagic and benthic prey whereas, in deeper waters, they fed predom- inantly on either benthic or pelagic sources depending on their habitat preferences. Our results support the hypothesis of a stronger benthic-pelagic coupling in shallow waters, notably through fish diet, and highlight the importance of including environmental factors such as depth as proxies of habitat variation to fully understand resource use and food web structure in epicontinental seas.
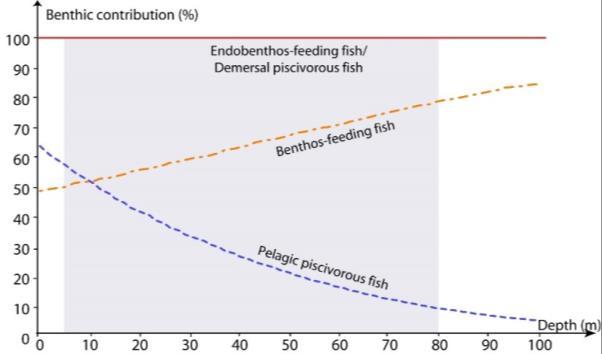
Figure 6 :Variation of the proportional benthic contribution (all benthic sources summed) to the diet of fish functional groups (proportional pelagic contribution is simply obtained as 1-benthic contribution). Blue dashed line: pelagic piscivorous fish; orange dash-dot line: benthos-feeding fish; red continuous line: demersal piscivorous and endobenthos-feeding fish. Giraldo et al. 2017.
Seasonal and ontogenetic variation of whiting diet in the Eastern English Channel and the Southern North Sea: An accurate description of trophic interactions is crucial to understand ecosystem functioning and sustainably manage marine ecosystems exploitation. Carbon and nitrogen stable isotopes were coupled with stomach content analyses to investigate whiting (Merlangius merlangus, Linnaeus, 1758) feeding behavior in the Eastern English Channel and Southern North Sea. Whiting juveniles and adults were sampled in autumn and winter to investigate both ontogenetic and seasonal changes. In addition, queen scallops (Aequipecten opercularis) samples were collected along with fish to be used as isotopic benthic baseline. Results indicated an ontogenetic diet change from crustaceans to fish and cephalopods. In autumn, ?15N values generally increased with fish size while in winter, a decrease of ?15N values with fish size was observed, as a potential result of spatial variation in baseline ?15N values. In winter, a nutrient-poor period, an increase in feeding intensity was observed, especially on the copepod Temora longicornis. This study provides further insights into whiting trophic ecology in relation to ontogenetic and seasonal variations, and it confirms the importance of combining several trophic analysis methods to understand ecosystem functioning (Figure7).
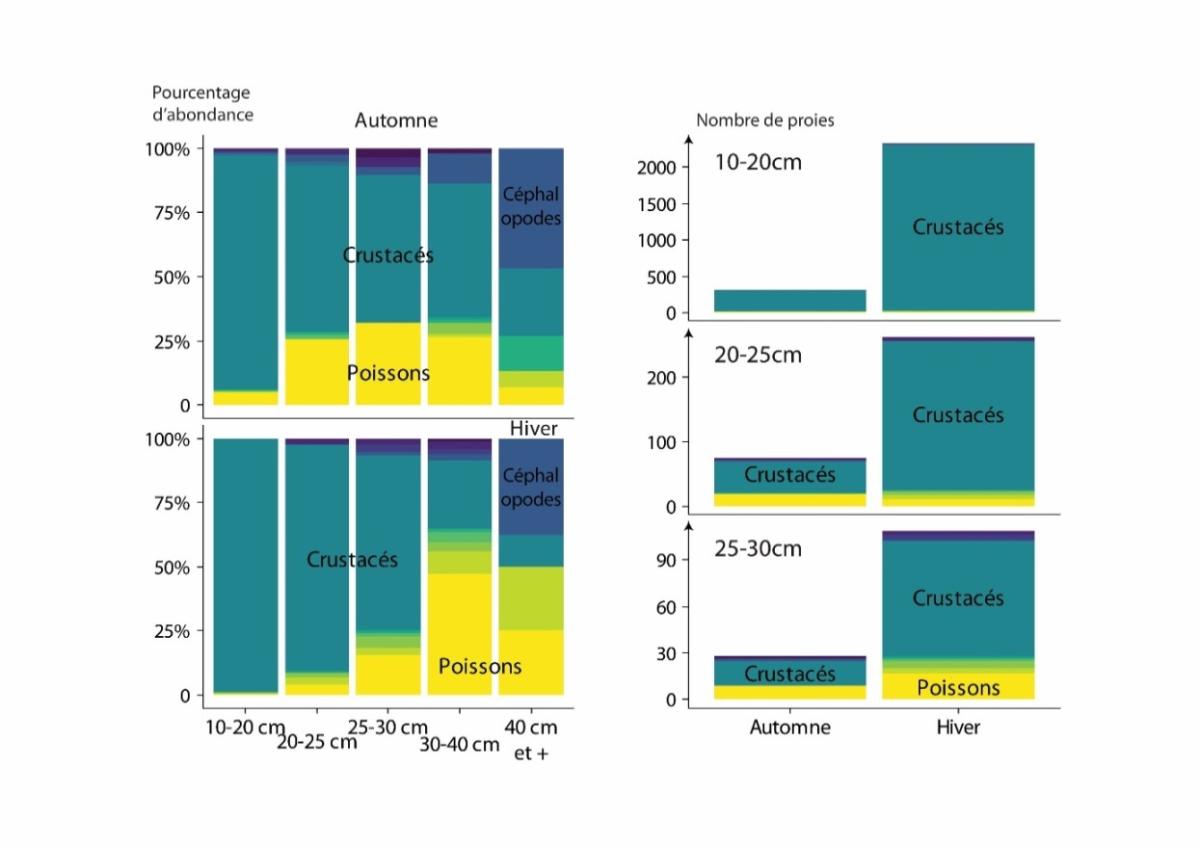
Figure 7: Ontogenic (Percentage of prey abundance in the diet, left) and seasonal (Total number of prey, by size class in fall and winter, right) changes in whiting diet. Redrawn from Timmerman et al. (2020).
Emergence of negative trophic level-size relationships from a size-based, individual-based multispecies fish model: Modeling the mechanisms underlying trophic interactions between individuals allows the food web structure to emerge from local interactions, which constitutes a prerequisite for assessing how marine ecosystems respond to various anthropogenic pressures. Using a multispecies spatially explicit individual-based model, the emergence of trophic patterns was explored in the eastern English Channel ecosystem, where pelagic-benthic trophic coupling was recently studied empirically. The OSMOSE model was applied to this ecosystem by explicitly representing the life cycle of 13 fish species and one squid group, forced by pelagic and benthic prey fields that are variable over time and space. A matrix defining possible accessibilities between life stages was added to the model to link benthic and pelagic communities through overlap of vertical distribution. After optimizing some parameters of the model to represent the average state of the fish community during the 2000¿2009 period, the simulated trophic structure was explored and compared to empirical data. The simulated and stable-isotope-derived trophic levels of fish were in relatively good agreement. Intraspecific variability of the trophic level is high in the five stable-isotope datasets but is well encompassed by the model. Despite the hypothesis of opportunistic size-based predation, the simulation showed a decreasing trend of trophic level with size for four benthic species, a pattern observed empirically for a different set of species in the ecosystem. Model exploration showed that this emerging pattern varies spatially and is both explained by the spatial variability of prey availability and by the independence of trophic and size structures of benthic invertebrates. The combination of individual-based models, stomach contents and intrinsic tracers, such as stable isotopes, appears to be a promising tool to better understand the causes of observed trophic patterns (Figure 8).
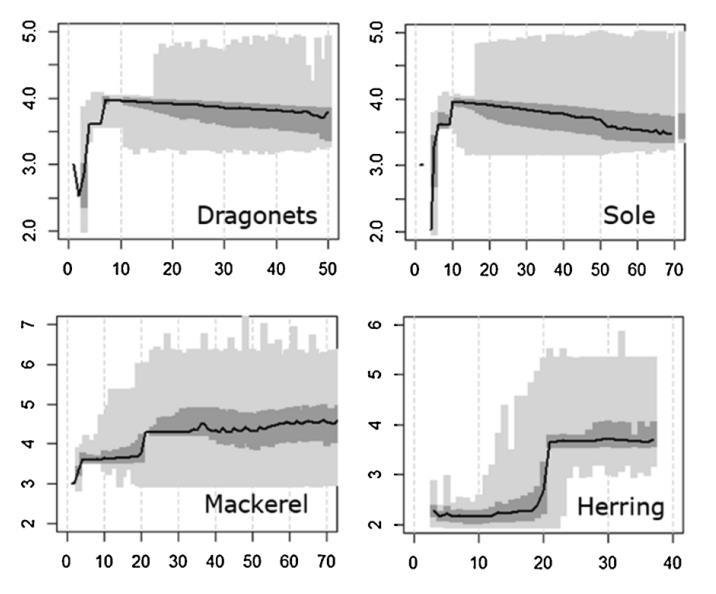
Figure 8 : Distribution of trophic levels predicted by the OSMOSE model for callionyma, sole, mackerel and herring. The increasing trends for mackerel and herring are consistent with the pelagic lifestyle of these two species, while the opposite trend for the other two species is consistent with their pelagic lifestyle (Travers-Trolet et al., 2019)
High frequency study of planktonic communities (M. Huret - IFREMER)
During the CGFS survey, the physico-chemical parameters of the water are analyzed at each haul using standard oceanographic protocols, to characterize the environment of the organisms studied and identify relationships between the environment and the distribution of these organisms. In addition to these abiotic parameters, the first links in the food chain, i.e. phytoplankton and zooplankton, are sampled and analyzed to know both their specific composition and their size structure. It is thus possible to identify a spatial structuring of planktonic communities, both from a taxonomic and functional point of view. The use of specific gear (bongo net and egg pump) makes it possible to locate and estimate the importance of spawning grounds and larval transitions areas, key habitats for fish populations. The samples collected are generally analyzed with a zoocam before being fixed and stored in vials. An image analysis is thus carried out on board in order to estimate the abundance of eggs and of the large zooplankton groups encountered before the end of the campaign (Figure 9).
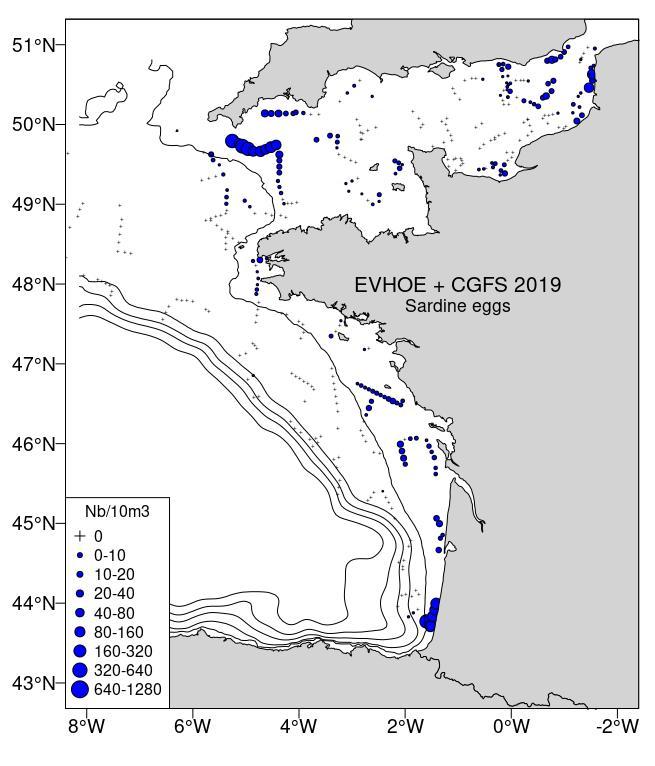
Figure 9: Sardine spawning habitat, by compiling data from CUFES + ZooCAM from CGFS and EVHOE survey
Functional Ecology¿¿
Ecological and life history traits explain a climate-induced shift in a temperate marine fish community: A better understanding of community dynamics and ecosystem functioning can be achieved by describing how community functional structure responds to environmental change over both time and space and by identifying which functional groups best mediate community responses. Here, we used a trait-based approach in combination with a newly developed application of principal response curves to functionally characterize a rapid taxonomic shift in the eastern English Channel fish community in the late 1990s. We identified the functional groups with the greatest contributions to the overall shift in fish functional structure and uncovered significant trait?environment relationships. We found that pelagic species with rapid life history cycles, characterized by broadcast spawning, small offspring size, and early maturation, declined considerably in abundance following an increase in sea surface temperature associated with a warming phase of the Atlantic Multidecadal Oscillation, which was likely exacerbated by historical fishing pressure. In contrast, species with late maturation, high parental care, and few, well-developed offspring increased in abundance, reinforcing that fish community responses to climate warming are strongly mediated through life history traits. By examining how environmental factors drove a community shift at the trait level, we provide a mechanistic understanding of how fish functional structure responds to rapid environmental change (Figure 10).
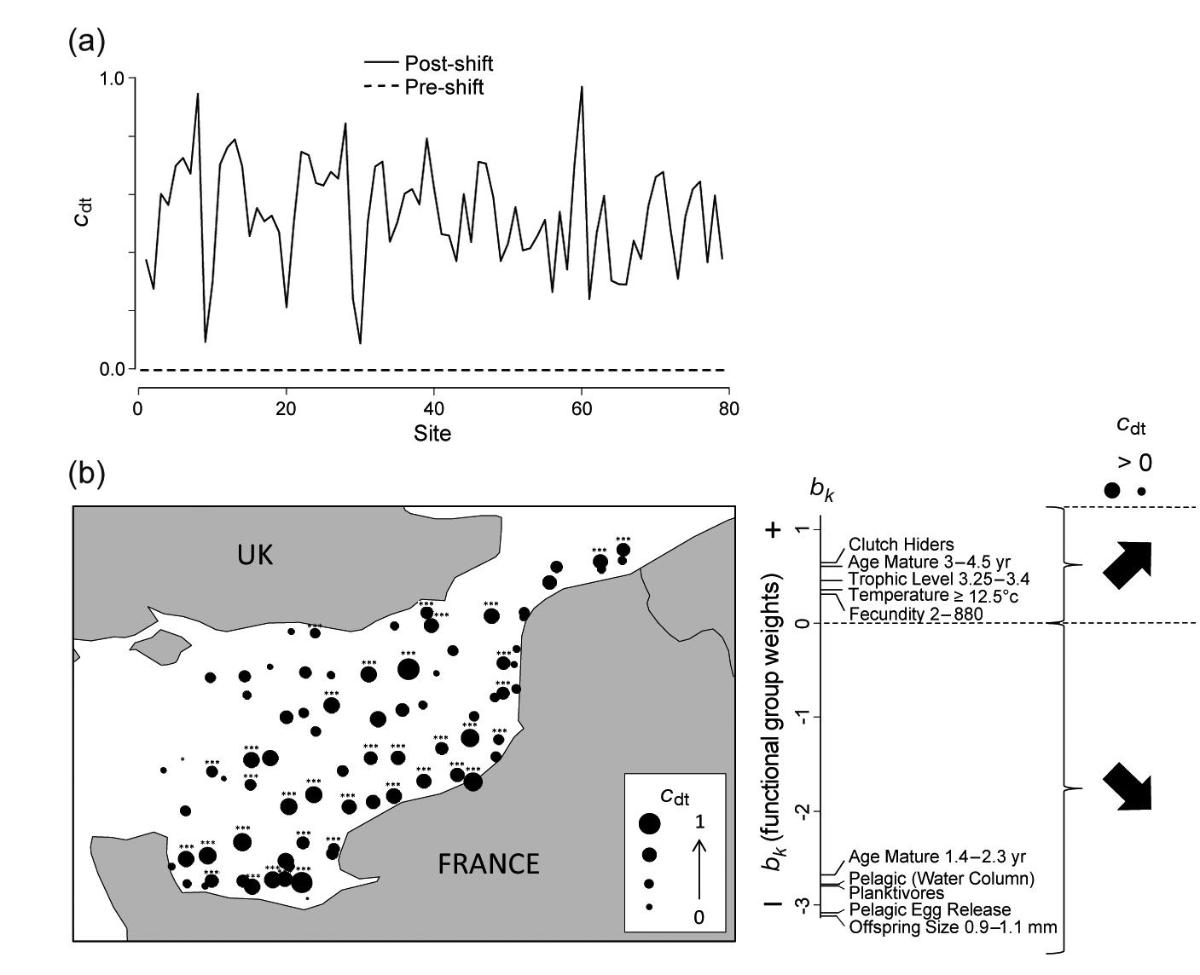
Figure 10: (a) Principal response curve showing changes in fish community structure across sampling sites (x-axis) between the baseline period (1988¿1997) (pre-shift) and tested period (1998¿2011) (post-shift). (b) Map showing the amplitude of temporal changes in fish functional structure (i.e. cdt values) at each sampling site and the trend (increase or decrease) of abundance for the most contributive functional groups (diagram to the right of map). Functional groups are ranked by their contribution weight (bk) coefficients. For clarity, only traits with bk coefficients in the first or last decile are shown. Sampling sites with significant change are shown by asterisks (***p < 0.001; **0.001 < p < 0.01; *0.01 < p < 0.05).
The RECCRU program (RECrutement CRUstacés) ( Morgane Guillam - Station Biologique de Roscoff)
RECCRU aims to set up recruitment indices for the main species of crustaceans of strong commercial interest from the Channel coasts such as lobster (Homarus gammarus), oil cake (Cancer pagurus), spider crab (Maja brachydactylus) and the red lobster (Palinarus elephas), in order to predict the evolution of their stocks and to put in place appropriate management measures. One of the axes of this program coordinated by the Roscoff Biological Station aims to acquire data on the larval phase of these commercial species with a particular interest for lobster. The characteristics of the early stages of planktonic life of these large crustaceans, such as the lifespan of larvae, their periods of presence, their spatial distribution as well as the potential links between larval abundances and benthic recruitment and connectivity between stocks, are the main aspects studied during this project.
In this context, as part of the CGFS surveys, a series of sampling has been carried out in the Western Channel for 3 years using a MIK plankton net in order to establish the spatial distribution of the larvae of the lobster Palinurus elephas in the Western Channel. To date, a network of 17 to 23 stations, i.e. a total of 61 stations, has been sampled since 2018. No lobster larvae have so far been collected during these campaigns. On the other hand, the preliminary results show the presence of phyllosoma larvae of sea cicadas (Figure 11) (morphologically similar to those of lobsters) mainly in the westernmost part of the Western Channel, west of the Ouessant front (Figure 12). We also note the presence of lobster and spider crab zoes. The year 2021 should end this spatio-temporal monitoring of crustacean larvae in the western Channel within the RECCRU project.

Figure 11: Phyllosome larva of Scyllarus arctus. The total length of the individual is 1.3cm.
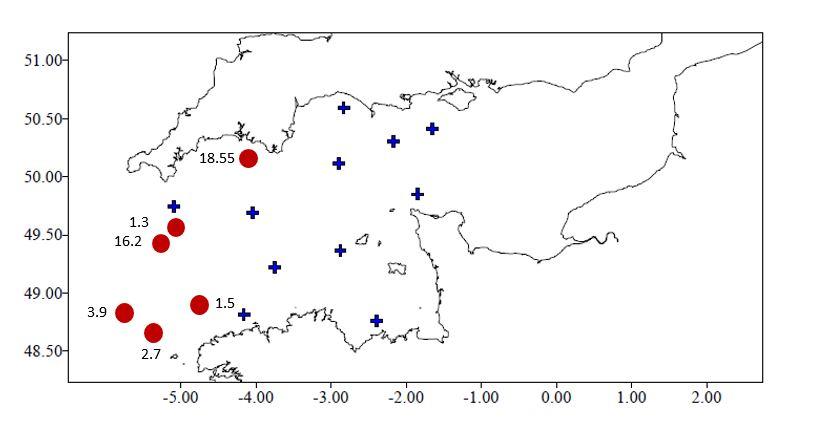
Figure 12: Spatial distribution of the phyllosoma larvae of Scyllarus arctus in the Western Channel, observed during the CGFS 2018 survey. The red dots indicate the presence of phyllosomas; larval abundance is expressed in larvae. 10000m-3. The blue crosses indicate the stations where no larvae were observed.

 , LE ROY Didier , MARTIN-BAILLET Victor
, LE ROY Didier , MARTIN-BAILLET Victor 