Pour les 3 espèces cibles retenues (i.e. les annélides polychètes Pectinaria koreni, Owenia fusiformis et Sabellaria alveolata), les abondances larvaires maximales ont été échantillonnées dans les plumes des estuaires de la Loire et de la Vilaine confirmant le rôle clef de tels structures sur le transport larvaire dans les environnements côtiers (Figure 1). Leur importance à agir comme des barrières physiques à un transport vers le large dépend cependant de leur variabilité temporelle au regard de la durée de vie des larves. Le long des côtes nord du Golfe de Gascogne, la variabilité saisonnière des conditions météorologiques favorise tantôt une expansion des plumes vers le nord-est et une connectivité entre populations côtières voisines, tantôt des upwellings côtiers et un export vers le large des eaux estuariennes.
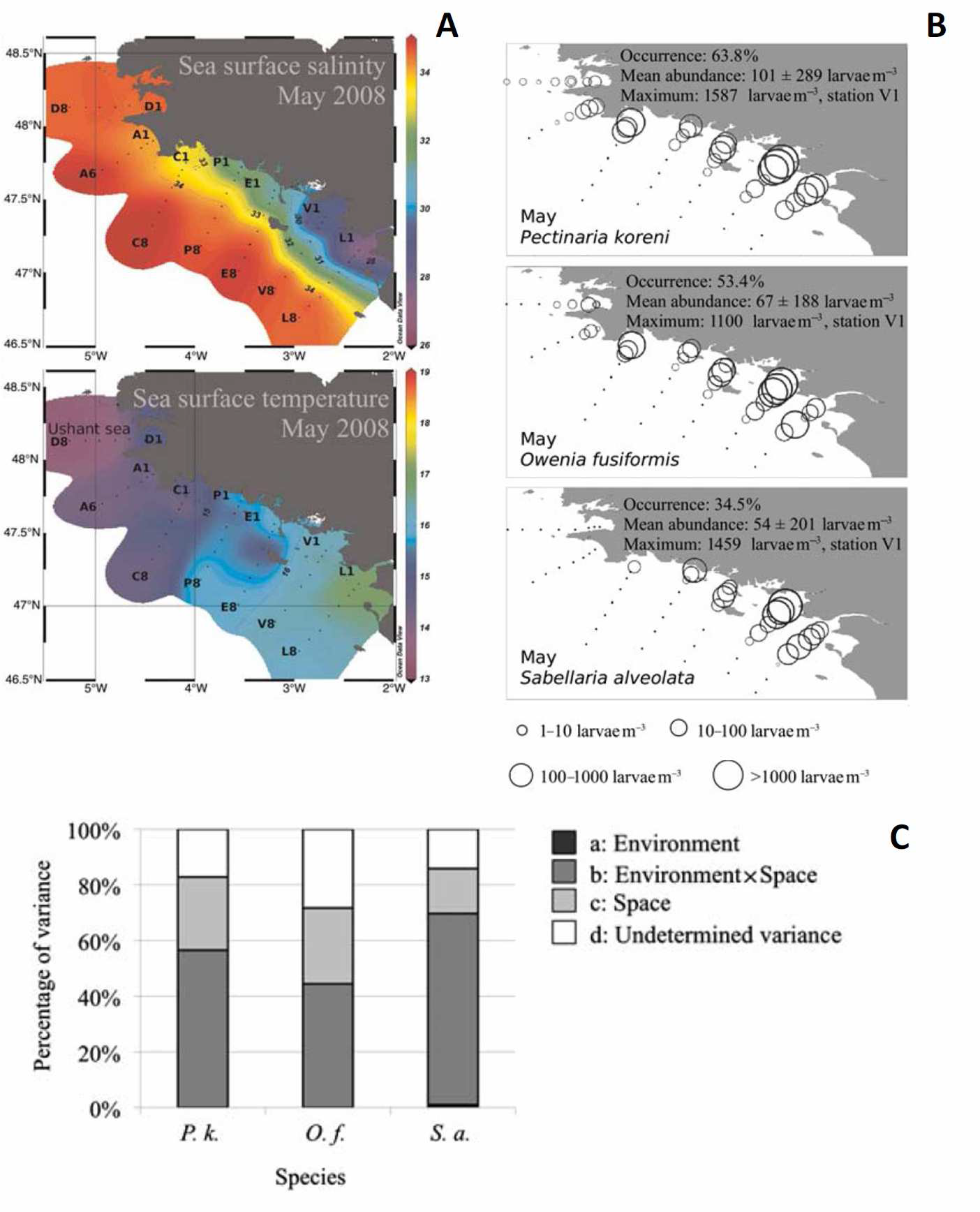
Figure 1. Structures hydrologiques et distributions larvaires dans le nord du Golfe de Gascogne en mai 2008. (A) Distribution de la salinité et de la température des eaux de surface. (B) Distribution des abondances larvaires (ind. m-3) de Pectinaria koreni, Owenia fusiformis et Sabellaria alveolata. (C) Partition de la variance des abondances larvaires totales de Pectinaria koreni (Pk), Owenia fusiformis (Of) et Sabellaria alveolata (Sa). D'après Ayata et al. (2011).
L'analyse de la distribution des larves a été réalisée à l'aide d'une méthode originale de partition de variance qui vise à différencier le rôle direct de l'environnement sur les réponses biologiques du rôle de l'organisation spatiale de cet environnement. En dépit d'une distribution des populations adultes et de traits d'histoire de vie variables selon les espèces étudiées, les patrons de distribution horizontale des larves des différentes espèces étaient expliqués par les mêmes variables hydrologiques (e.g. température de surface, salinité de surface, degré de stratification de la colonne d'eau). Cependant, si ces dernières peuvent agir directement sur le développement et la survie des larves, leur influence directe sur la variabilité des abondances larvaires est négligeable et n'excède pas quelques pourcents (Figure 1). C'est au contraire l'organisation spatiale de l'hydrologie, reflet de l'hydrodynamisme sur les caractéristiques des masses d'eau, qui explique l'essentiel de la variabilité des abondances larvaires, de 40 à 68 % de la variance totale. L'espace seul explique 10 à 34 % de la variance des abondances qui peuvent être attribués à des caractéristiques des espèces spatialement structurées : (i) distribution et taille des populations adultes, (ii) date et intensité des émissions de larves, (iii) sédentarisation des recrues. Enfin, en dépit de l'importance de l'hydrodynamisme, une proportion non négligeable de la variance des abondances larvaires, comprise entre 14 et 47 %, demeurait inexpliquée. Deux processus majeurs peuvent être responsables de cette part inexpliquée : (i) la nature stochastique de la circulation côtière induisant une distribution agrégative des larves à petite échelle et (ii) les comportements larvaires à même de modifier la distribution horizontale des larves comme cela a été rapporté pour les 3 espèces.
Les patrons observés de distribution des larves sont voisins des patrons simulés avec des larves dont le transport dépend largement de la dynamique des plumes estuariennes en complément à différents facteurs biologiques : localisation des populations adultes, date de ponte, durée de vie larvaire et comportement migratoire (Ayata et al., 2010).
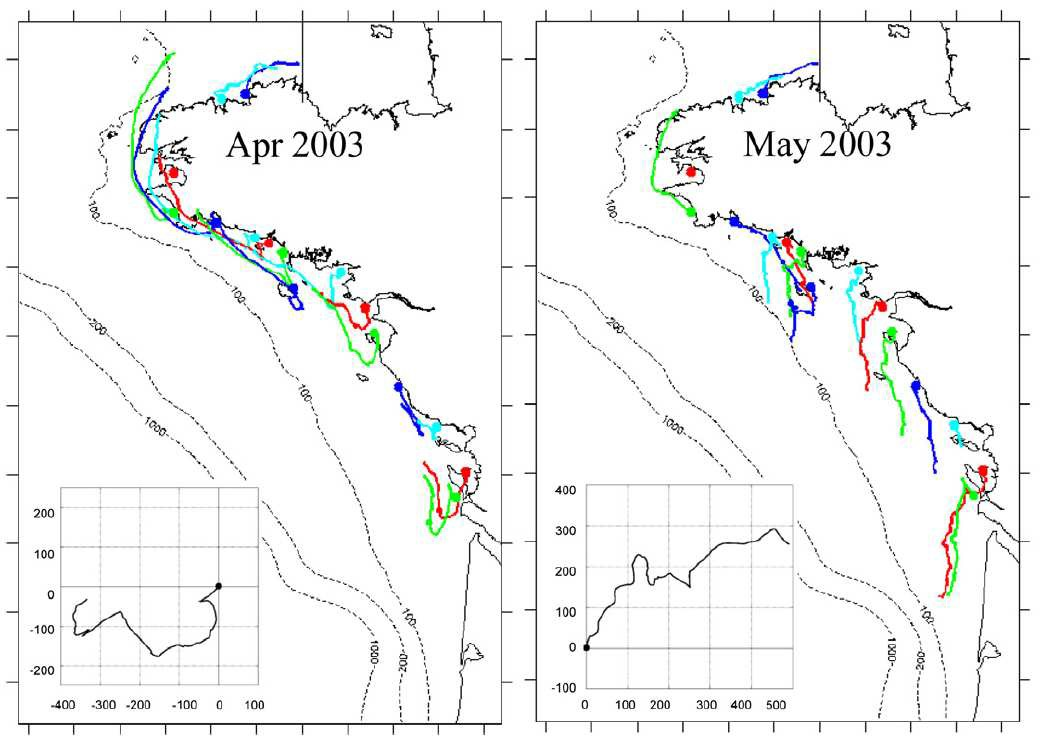
Figure 2. Trajectoires moyennes simulées de particules émises en avril et mai en fonction de la localisation de la population génitrice. Les larves étaient considérées passives. La durée de vie larvaire était fixée à 4 semaines. Les conditions de vent pendant la durée des simulations sont figurées pour chaque mois sous forme d'un "progressive vector diagram". D'après Ayata et al. (2010).


