171 opérations (dragages et chalutages) entre 100 et 1000 mètres de profondeur ont été réalisées. Les résultats de la campagne peuvent être regroupés en trois ensembles :
- Des résultats touchant à l'inventaire et à la description de la faune, notamment des descriptions d'espèces nouvelles ;
- Des résultats touchant à la biogéographie, en particulier l'étude de l'isolement et de l'endémisme ;
- Enfin, des résultats d'opportunité, fondés pour partie sur des échantillons prélevés au cours de la campagne EBISCO, mais qui auraient pu l'être ailleurs.
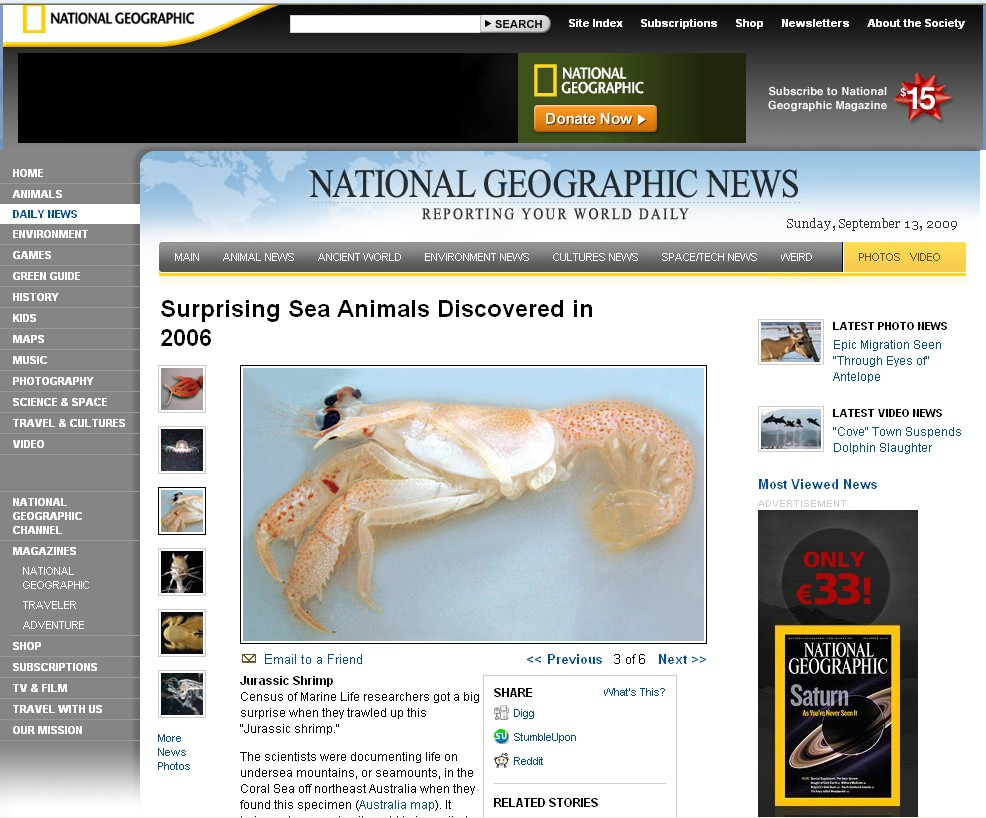
La découverte de Neoglyphea neocaledonica sur le Banc Capel, par 367-536 m de profondeur, illustre à la perfection la nature imprédictible des résultats des campagnes d'exploration. En effet, la campagne EBISCO a eu la chance de recueillir - de manière évidemment entièrement fortuite - un spécimen d'une espèce nouvelle de crustacé glyphéide - la deuxième espèce du groupe. La première espèce avait été pêchée en 1908 aux Philippines par un navire américain mais l'unique spécimen était resté non identifié jusqu'à ce que Michèle de Saint Laurent et Jacques Forest publient sa description en 1975 sous le nom de Neoglyphea inopinata. La découverte d'un spécimen vivant du groupe des glyphéides, que l'on croyait éteint depuis la fin de l'ère secondaire, avait alors déjà fait grand bruit: « One of the more interesting crustacean discoveries of the last century was the recognition and description of Neoglyphea inopinata Forest & de Saint-Laurent, 1975, a living member of a group long thought to be extinct since the Mesozoic » (Schram & Ahyong, 2002).
La découverte de ce spécimen vivant permet évidemment l'utilisation des outils moléculaires. L'hypothèse actuelle la mieux soutenue, à la fois par les études morphologiques et moléculaires, est que les glyphéides sont proches des Astacidea, alors que la morphologie des pattes (P1) est proche de celle des Achelata. L'absence de vraie pince chez les glyphéides n'aurait donc pas la même origine que chez les Achelata et il s'agirait alors d'une convergence fonctionnelle. Les Thalassinidea, chez lesquels ont observé également des pseudo-pinces correspondant à des adaptations à la vie en terriers, sont également éloignés des Astacidea. Le séquençage de Neoglyphea inopinata et de Laurentaeglyphea neocaledonica doit permettre d'étayer ou d'infirmer ces hypothèses en améliorant la qualité des relations phylogénétiques pouvant être établies par l'association de séquences sur 6 gènes : 3 gènes mitochondriaux et 3 gènes nucléaires.
Au cours de la campagne EBISCO, Bertrand Richer de Forges a immédiatement vu à bord du navire la découverte qui venait d'être faite, mais malgré plusieurs coups de drague sur la même position, aucun autre spécimen n'a été collecté. De la récolte en octobre 2005 à la description formelle en mars 2006, il s'est passé moins de 6 mois. La même année, Jacques Forest, le "père" des glyphéides actuels considérait que des différences importantes distinguent Neoglyphea neocaledonica de N. inopinata, et établissait pour elle le nouveau genre Laurentaeglyphea, qu'il jugeait d'ailleurs "beaucoup plus proche des formes fossiles".
Ce résultat sensationnel a donné lieu à la publication d'un communiqué de presse qui a été très largement repris et diffusé, depuis le site du Ministère de l'Enseignement Supérieur et de la Recherche (http://www.science.gouv.fr/fr/actualites/bdd/res/2309/t/4/decouverte-d-un-crustace-fossile-vivant-en-mer-du-corail/) jusqu'à celui National Geographic (http://news.nationalgeographic.com/news/2006/12/photogalleries/sea-animals/photo3.html, ci-dessous). Neoglyphea neocaledonica a également été retenu dans le "Top 10" des découvertes 2006 du Census of Marine Life. Une recherche sur Google avec les mots clés Neoglyphea neocaledonica donne encore (12 sept. 2009) 1270 hits.
Un autre résultat de la campagne EBISCO concerne la mise en évidence d'un endémisme très significatif sur les bancs et monts-sous-marins de la Mer du Corail. Quand nous avons démarré l'exploration du benthos profond en Nouvelle-Calédonie dans les années 1980, tout paraissait nouveau, mais nous nous attendions à ce que ces nouvelles espèces se retrouvent dans les archipels voisins quand leur tour viendrait d'être exploré. Vingt ans plus tard, ces autres archipels ont maintenant été couverts mais, à notre grand étonnement, la Nouvelle-Calédonie ressort de cette exploration comparative comme un endroit particulièrement original et riche. Aucune autre grande île du Pacifique Sud n'a des fonds durs aussi étendus. De plus, la ZEE de Nouvelle-Calédonie est topographiquement très hétérogène, ce qui contribue sans aucun doute aux divers niveaux d'endémisme régional emboités les uns dans les autres.
Plusieurs zones d'endémisme local peuvent être reconnues. La Mer du Corail a ses propres espèces endémiques (Perotrochus deforgesi, Calliostoma chesterfieldense, Cassis abbotti, Xastilia kosugei, Murexsul metivieri, Amalda coriolis, Leiosyrinx liphaima, Conus estivali), certaines paraissant même être restreintes de façon significative à un seul banc ou plateau, comme dans le cas des olividés des genres Belloliva et Calyptoliva (ci-dessous).
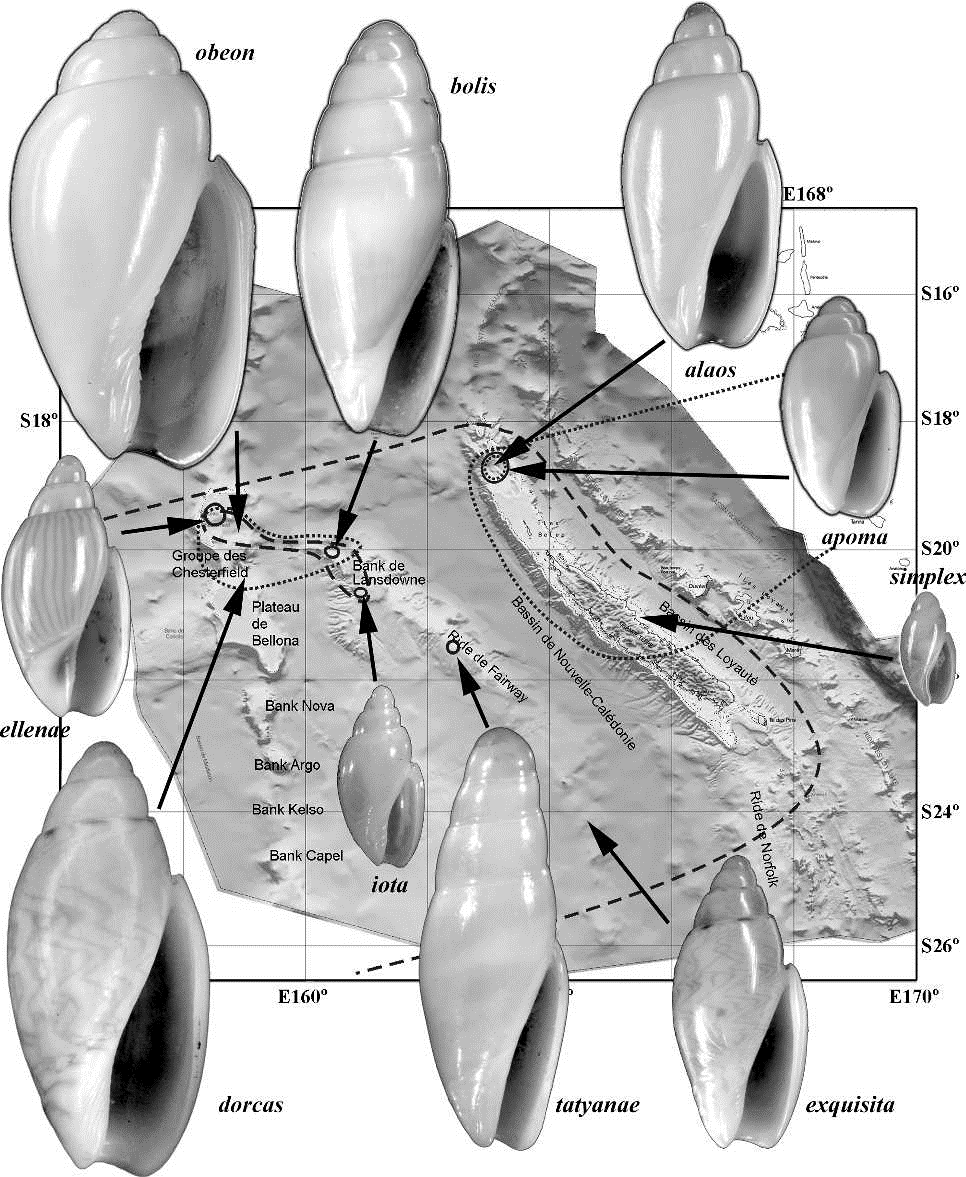
Les genres Belloliva et Calyptoliva (Gasteropoda, Olividae) montrent une splendide radiation en Mer du Corail et dans le Nord de la Nouvelle-Calédonie, avec 5 espèces micro-endémiques sur le plateau des Chesterfield et le Banc Lansdowne-Fairway. (Tiré de Kantor & Bouchet 2007).
Enfin, on peut faire état de résultats que l'on peut qualifier d'opportunité, car fondés pour partie sur des échantillons prélevés au cours de la campagne EBISCO, mais qui auraient pu l'être ailleurs. Entrent dans cette catégorie, les travaux de Nicolas Puillandre sur la phylogénie moléculaire des gastéropodes. Sa thèse, soutenue en septembre 2008, s'appuie sur un cocktail d'échantillons provenant de diverses campagnes et expéditions conduites de 2004 à 2007, dont EBISCO.

