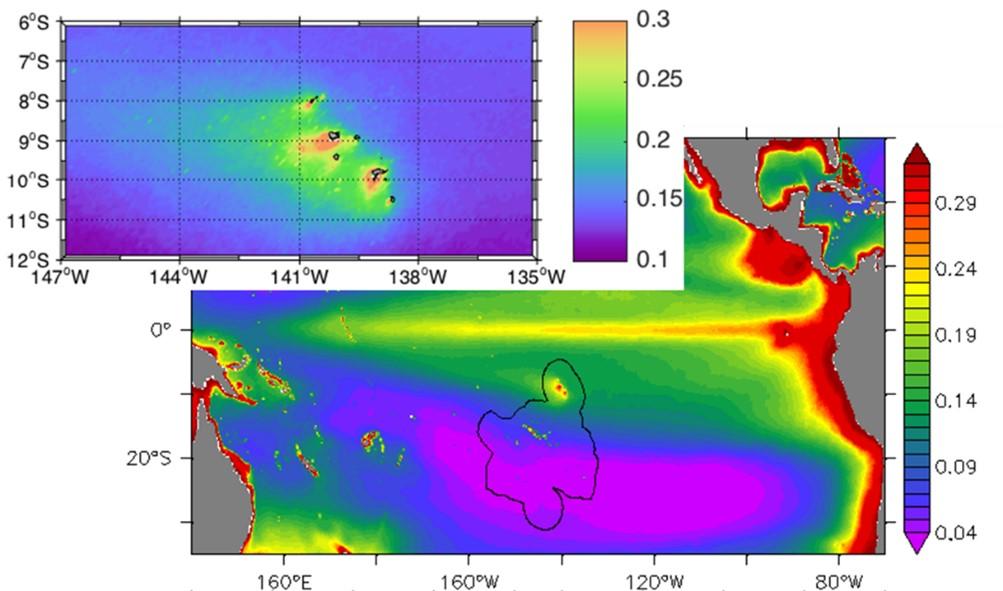
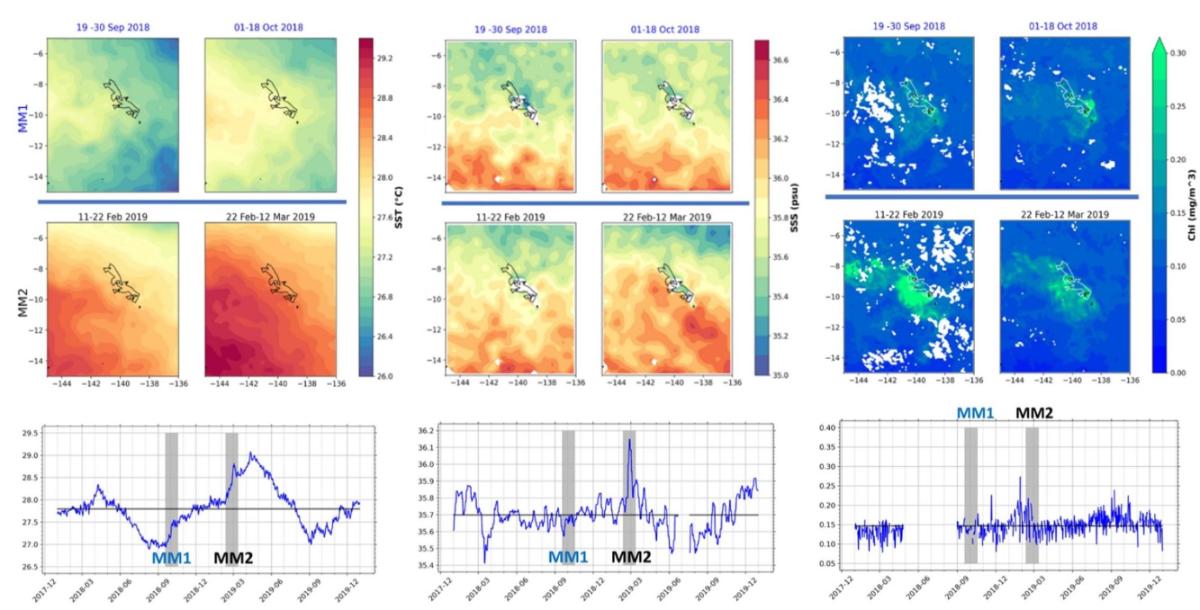
Circulation océanique de surface, énergie cinétique et turbulente : données S-ADCP et bouées dérivantes SVP (Le Roux C. 2020 ; Castant J., 2020 ; Martinez et al., 2022 - com. orale Maes C.)
Les mesures obtenues à partir du S-ADCP (Ship borne Acoustic Doppler Current Profiler) du NO Alis ont permis d'établir une cartographie des courants au cours la campagne (Fig. 4). En été austral l'influence du SEC semble bien prédominante avec des courants globalement orientés vers Ouest (NO et SO), contrastant avec la situation observée en hiver pendant MOANA-MATY 2018 avec des courants de surface très variables (Rodier, 2018)
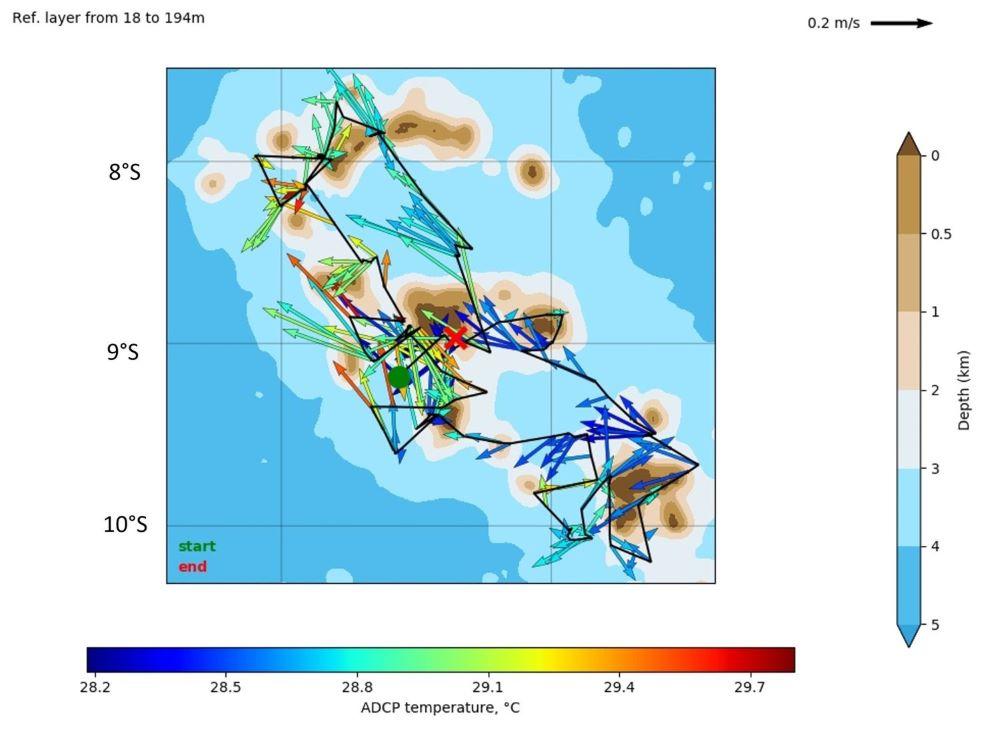
Figure 4. Courants moyens enregistrés au cours de la campagne MOANA-MATY 2 dans la couche 18-194m.
En complément des données ADCP, l'analyse des données dérivées des bouées dérivantes SVP et en particulier celles déployées lors des campagnes MOANA-MATY près des îles, a permis d'établir une climatologie saisonnière des courants de surface, permettant de replacer les campagnes dans un contexte plus général. Plus de 1000 jours-bouées ont ainsi été recensées entre 2018 et 2019 dans la région des Marquises dont environ 95 jours-bouées par cellule 1°x1° entre Janvier et Mars 2019 dans la zone étudiée pendant MOANA-MATY 2019 (9°S-8°S, 144°W-141°W).
Les bouées SVP déployées pendant les campagnes 2018 et 2019 montrent des trajectoires très différentes (Fig. 5A), reflétant la variabilité saisonnière des courants de surface dans zone des Marquises (Fig.5B). A plus fine échelle, ces données SVP permettent d'estimer les flux inter-îles ainsi que leur variabilité saisonnière. Par exemple, en été comme pendant MOANA-MATY 2019, le courant qui longe la côte nord de Hiva Oa est très faible (6.5 cm.s-1) et s'écoule vers le SO, ce qui est différent en hiver. Entre Ua Huka et Hiva Oa, le courant est SO et plus fort en été. Entre Ua Pou et Nuku Hiva et au nord de Nuku Hiva, le courant est aussi plus fort en été (23 cm.s-1), et faiblit de Juillet à Décembre (12 à 14 cm.s-1).
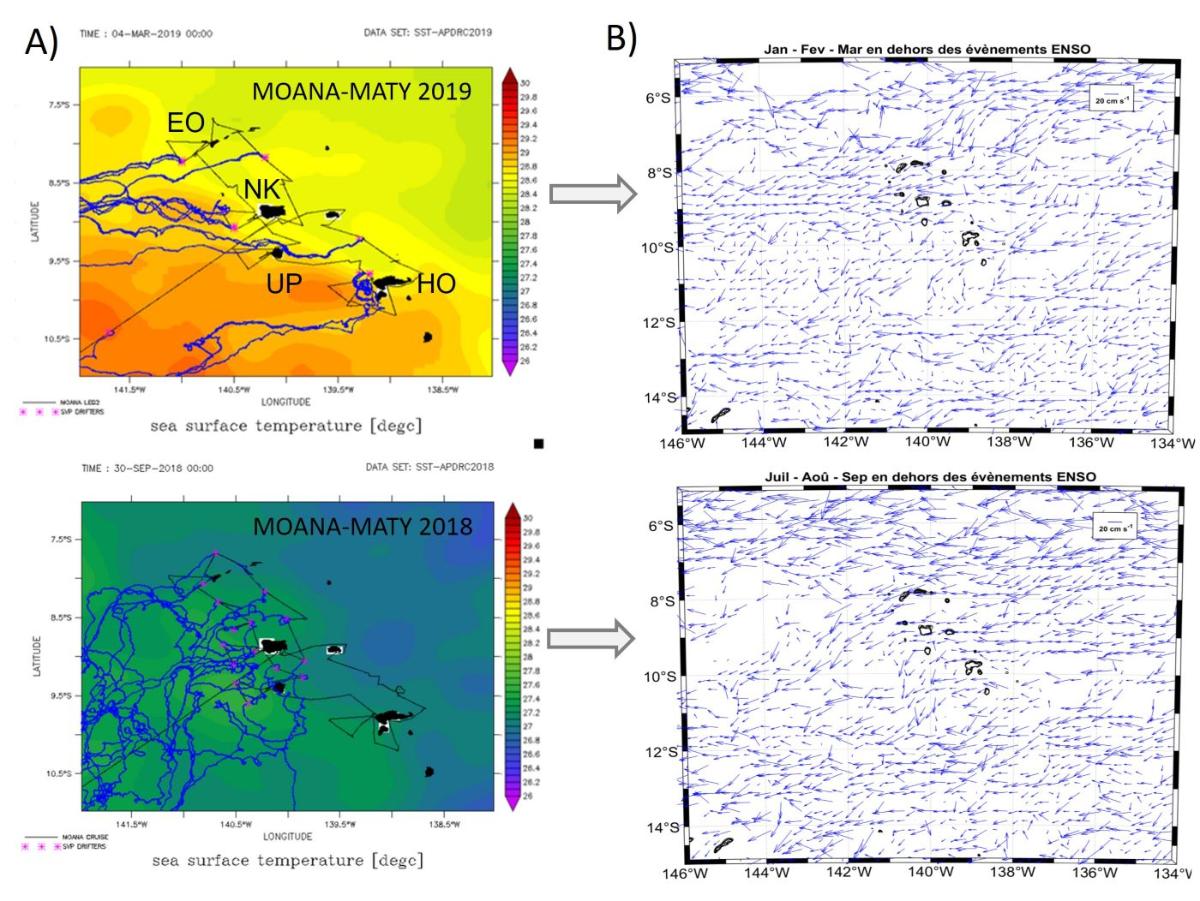
Figure 5. A) Trajectoires des bouées dérivantes SVP déployées pendant MOANA-MATY 2018 et 2019; B) Climatologie saisonnière des courants de surface sur la période 1979-2019 calculée à partir des données issues des bouées SVP (incluant celles déployées pendant les campagnes). Résolution spatiale de 1/4 de degré. Seuls sont présentés les résultats en été et hiver, correspondants aux deux MOANA-MATY.
Le traitement des données SVP a également permis de définir certaines caractéristiques liées à l'effet d'île, telles que l'énergie cinétique (MKE) et turbulente (EKE). L'énergie cinétique (MKE) au niveau des Marquises varient saisonnièrement comme la courantologie (Fig. 6A), de 120-175 cm2.s-2 en été austral (MOANA-MATY 2019) à 150-180 cm2.s-2 à la fin de l'hiver (MOANA-MATY 2018). Elle est toujours plus faible en aval de l'île de Hiva Oa qu'en amont et plus élevée au nord de l'archipel qu'au sud, avec des maxima au nord de 7°S en été (> 400), caractéristiques de la zone équatoriale. L'étude du rapport EKE/MKE (Fig. 6B) permet d'identifier les zones où la turbulence est prédominante, notamment dans la partie sud de l'archipel et en aval des îles (O-SO). L'archipel des Marquises se trouve toutefois dans une région où le rapport reste relativement faible (< 2 en moyenne, maximum 5.4 en été au SO de Nuku Hiva), comparé aux valeurs beaucoup plus sud (30 à 100, au sud de 30°S).

Figure 6. A) Energie cinétique moyenne MKE et B) ratio EKE/MKE sur la période 1979-2019, calculés à partir des données SVP. Résultats en été (haut) et hiver (bas), correspondants aux 2 campagnes MOANA-MATY 2019 et 2018.
Ces résultats sur la dynamique océanique (ADCP, SVP) seront complétés par les données issues de SPASSO « Software Package for Assimilation of Satellite data in Oceanography». Des cartes basées les diagnostics lagrangiens sont déjà disponibles sur le site SPASSO (https://spasso.mio.osupytheas.fr/MOANA_MATY/Figures_web/), mais il faudra étendre ces expériences lagrangiennes (étude des processus à fine échelle, tourbillons, ondes internes) afin de définir l'origine et le devenir des enrichissements et les liens avec les îles (coll. S. Barillon, A. Petrenko, MIO).
Biomasse, production et composition des communautés phytoplanctoniques (Charavit J., 2019 ; Martinez et al., 2022 - com. orale Rodier M.; Layec et al., 2024)
Biomasse et production phytoplanctonique (Fig. 7). Lors de la campagne MOANA-MATY 2019, des enrichissements de surface en Chla ont été observés en aval des îles, particulièrement au sud-ouest de Nuku Hiva, atteignant 1 mg.m-3 dans les 30 premiers mètres. À l'inverse, les zones situées en amont des îles présentent un faible enrichissement, avec un enfoncement du maximum de chlorophylle (DCM), (> 60 mètres, et jusqu'à 106 mètres à la St. 6). Cette situation diffère de celle observée pendant MOANA-MATY 2018 où le contraste aval/amont était peu marqué, ce qui peut être lié à la variabilité saisonnière des régimes de courants (Fig. 5).
La biomasse en Chla est largement dominée par le picoplancton (< 2 µm) et les picocyanobactéries, sauf dans les zones enrichies au SO de Nuku Hiva, où 50 % de la Chla est constituée d'organismes > 10 µm, associés au développement de diatomées (cf. & suivant). Au large de Hiva Oa, l'enrichissement semble plutôt lié à des organismes plus petits, pico- et nanoplanctoniques (incluant des dinoflagellés), ce qui suggère des stades d'évolution différents de l'écosystème.
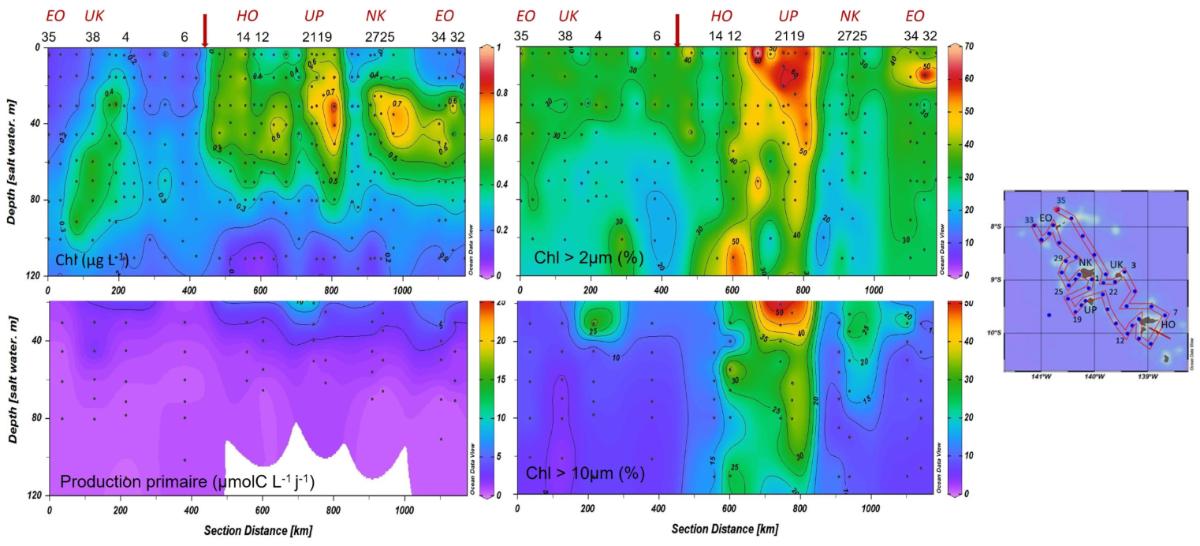
Figure 7. Distribution de la Chla totale et fractionnée (>2 et 10µm)) et de la production primaire au cours de la campagne MOANA-MATY 2019. Distance le long du trajet indiqué sur la carte. EO (Eiao), UK (Ua Huka), HO (Hiva Hoa), UP (Ua Pou), NK (Nuku Hiva).
Les valeurs intégrées de Chla sur la couche euphotique (0,1% de lumière) varient entre 18,1 et 52,7 mg Chl-a m-2 (33,1 ± 9,0 mg Chla m-2) et sont associées à des valeurs de production primaire variant de 56 à 433 mg C m-2 j-1 (219 ± 103 mg C m-2 j-1) et maximales en surface (0-20/30m). Ces valeurs confirment le caractère mésotrophe (type HNLC) des eaux océaniques de l'archipel des Marquises, déjà observé lors de MOANA-MATY 2018. Toutefois, contrairement à la campagne de 2018, la production phytoplanctonique comme la biomasse se concentrent essentiellement sous le vent des îles (Fig. 7), ce qui peut s'expliquer en partie par des différences dans les régimes de courants. L'écosystème pélagique autour de l'archipel des Marquises apparaît donc comme un système globalement productif de type HNLC, mais aussi très dynamique, comme le montrent les images satellites prises lors de la campagne (Fig.8).
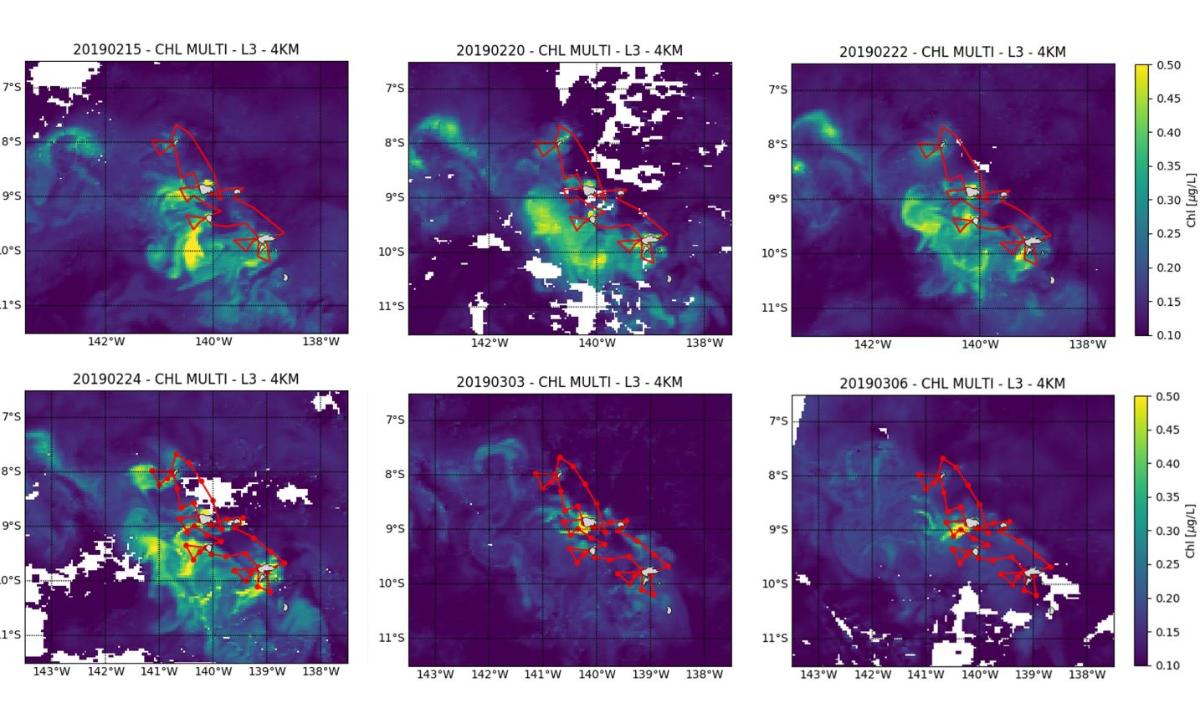
Figure 8. Carte de Chlorophylle (µg L-1), dérivée des données satellites GlobColour (MERIS/MODIS/VIIRSN) au cours de la campagne MOANA-MATY 2019.
Composition des communautés nano-microplanctoniques. La composition du nano- et microphytoplancton a été analysée par microscopie optique couplée à une approche métagénomique aux stations longues. Les données pigmentaires par HPLC complèteront cette étude quantitative sur la structure des communautés phytoplanctoniques.
a) Les observations en microscopie mettent en évidence des changements de communautés au sein de l'archipel, associés à une grande variabilité en termes d'abondance (Fig. 9). Les flagellés dominent en diversité, représentés par deux groupes principaux : les Prymnésiophytes (16 espèces/genres) avec une dominance du genre Gephyrocapsa, et les Dinoflagellés (78 espèces). Les diatomées (64 espèces) sont particulièrement abondantes dans les zones riches situées en aval des îles avec de fortes valeurs de surface aux Sts 19 et 25, situées respectivement à 85 et 55km des îles.
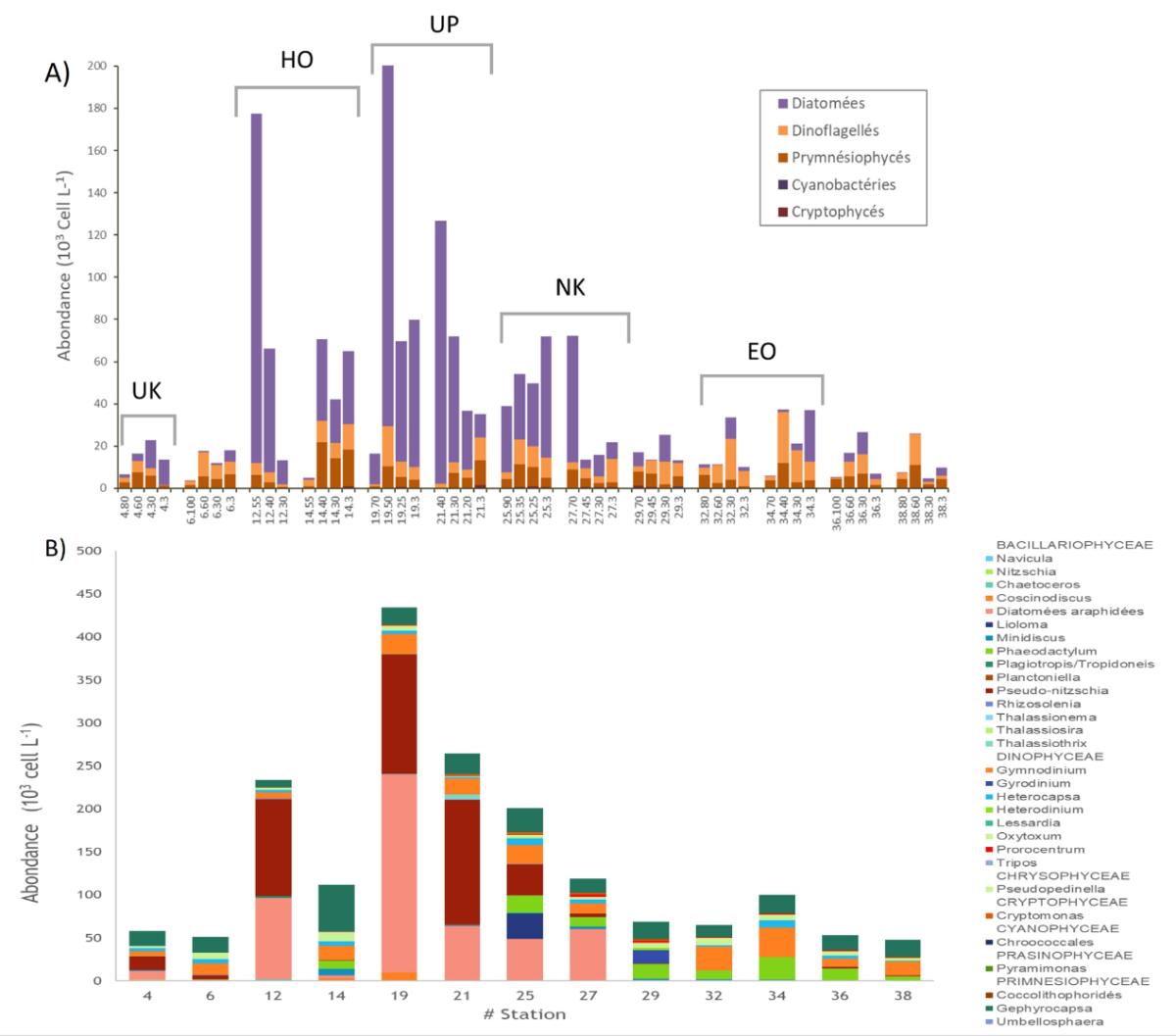
Figure 9. A) Distribution des principaux groupes taxonomiques sur l'ensemble de la couche euphotique pendant MOANA-MATY 2019 (observations au microscope inversé). Autour des îles, les stations les plus côtières sont à droite dans chaque encadré. B) Principaux genres présents.
b) L'ajout de données de metabarcoding pendant MOANA-MATY 2019 a permis d'obtenir une description plus détaillée de la structure et composition des communautés en surface et dans le DCM. Une première analyse des données a été réalisée sur les données > 3µm en identifiant trois clusters (Upstream (U) = aval, Côtier (C), Dowstream (D) = amont), une classification susceptible d'évoluer. Les analyses reposent sur les abondances relatives des séquences d'ADN obtenues après séquençage, exprimées en « reads » ou ASVs après traitement bioinformatique.
A l'échelle de l'archipel, les dinoflagellés dominent avec jusqu'à 50% des "reads", suivis des diatomées à hauteur de 30%, en accord avec les observations en microscopie. En surface, les stations situées en amont des îles (Sts 34, 36, 38 et 6) présentent les richesses spécifiques les plus élevées, entre 700 et 800 ASVs (Fig. 10). Les autres stations ont des richesses globalement plus faibles, avec des minima aux Sts 14 et M12 et très peu d'ASVs partagés avec les autres stations. Les diatomées sont le seul groupe à présenter en surface une corrélation positive entre l'abondance de "reads" et la distance à la côte (modèle linéaire, p-value < 0.05).
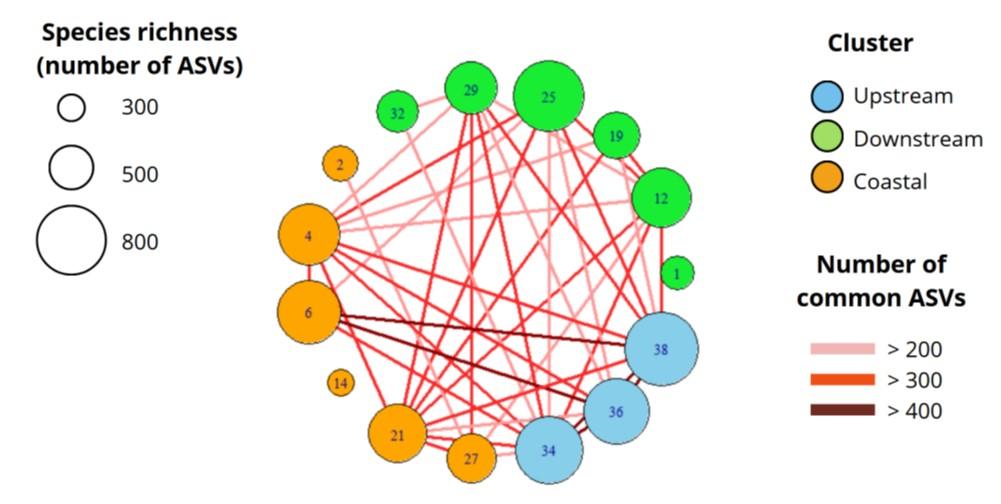
Figure 10. Richesse spécifique en surface par station et nombre d'ASVs communs. 1=M1.
Les données de métabarcoding et de microscopie montrent une relation inverse entre la richesse et diversité spécifique (indice de Shannon) et la concentration en Chla (p-value < 0,05), en accord avec les modèles théoriques. Cependant, aucun modèle distinct de type "courbe en dos de chameau" (Torok et al., 2016) ne se dégage de l'ensemble des données. Les indices de diversité varient selon la méthode d'analyse, mais en se basant uniquement sur les données de métabarcoding, on observe que les stations situées en amont présentent une diversité d'espèces plus élevée (Fig. 11A). A l'inverse, les diversités les plus faibles sont observées aux stations côtières Sts 14 et M1 situées en aval des îles.
L'analyse par NMDS/Dissimilarité de Bray-Curtis a permis de visualiser et de quantifier les variations de composition spécifique entre les stations (Fig. 11B). Les stations situées en amont des îles (U), y compris la St. 6, montrent une composition relativement homogène. En revanche, le groupe aval (D) présente une certaine hétérogénéité, avec une nette opposition entre les stations nord (29, 32) et les stations sud (12, 25 et 19). Enfin, les stations côtières (C) présentent une forte variabilité, avec la St. 14 (nord de Hiva Oa) qui se distingue encore des autres stations. Une relation positive significative a été observée entre la distance géographique (distance à la côte) et la dissimilarité de Bray-Curtis entre stations (statistique de Mantel = 0,26, p-valeur < 0,05).
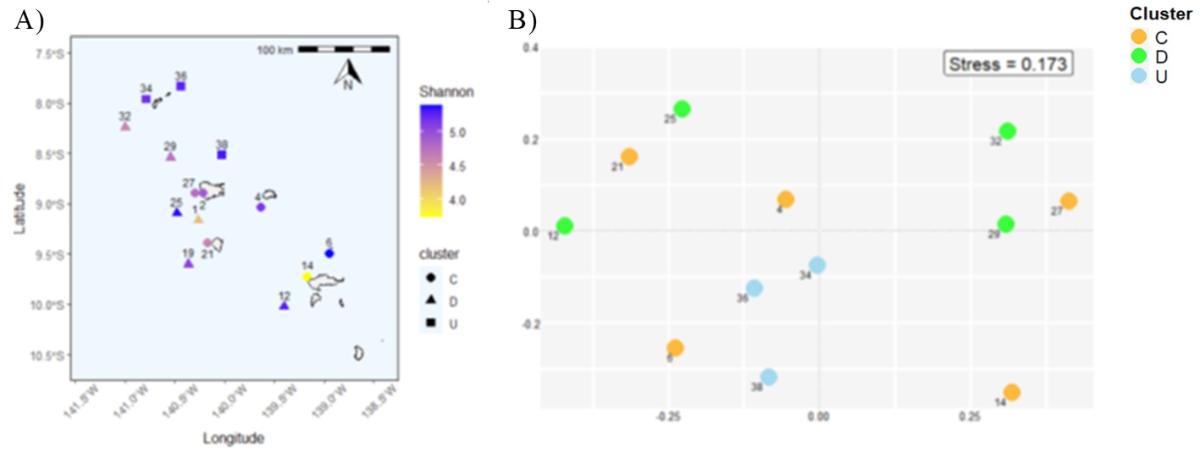
Figure 11. A) Distribution de l'indice de Shannon ; B) analyse NMDS utilisant la dissimilarité de Bray-Curtis calculée à partir du nombre relatif de "reads" par échantillon (D= dowstream, C=côtière ; U=upstream). Données de métabarcoding.
Les données de surface ont été comparées à celles dans le DCM, en ciblant les espèces présentes dans les deux couches (Fig. 12). Les résultats montrent un nombre de "reads" plus élevé en surface. L'absence de corrélation significative entre les rangs des ASVs en surface et dans le DCM souligne des différences marquées dans la structure des communautés entre ces deux niveaux, résultat également confirmé par les comptages par microscopie. Par exemple, à la station St. 14, une forte dominance de Thalassiosira tenera est observée en surface entraînant une faible diversité (Fig. 11), alors que la diatomée benthique Cylindrotheca closterium prédomine dans le DCM. Ces résultats suggère que les enrichissements observés en surface dans le panache des îles ne proviennent pas d'une advection passive d'organismes par mélange vertical («apparent bloom» ; Hasegawa et al., 2009), mais plutôt liés à des processus de fertilisation en surface.
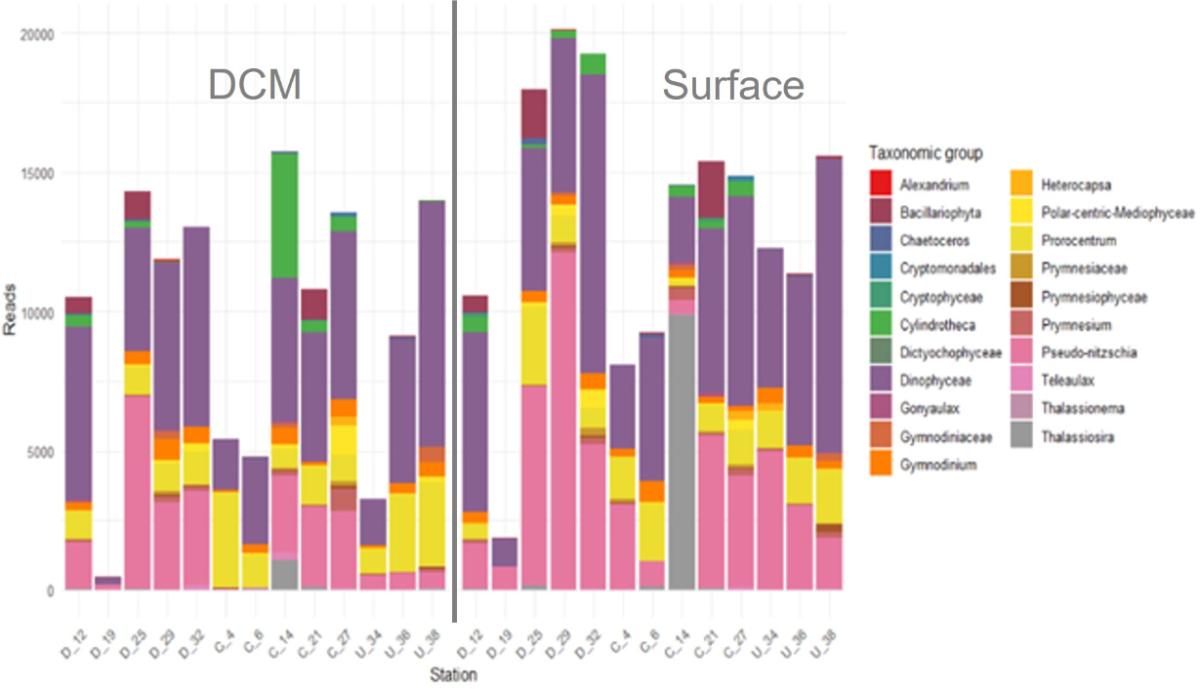
Figure 12. Principaux taxa communs de nano- et microphytoplancton (> 3µm) entre la surface et le DCM (de la classe au genre); données de métabarcoding.
Les données de microscopie et de métagénomie ont mis en évidence des différences dans la structure des communautés phytoplanctoniques entre les zones amont/aval et nord/sud de l'archipel. En amont, les communautés semblent relativement stables (forte similarité en termes de richesse et diversité spécifique entre stations), et adaptées aux conditions mésotrophes de la zone. En aval, dans le sillage des îles, la richesse spécifique tend à diminuer tandis que la biomasse et production augmentent ; la stabilité est détruite et une forte hétérogénéité entre station apparait au niveau de la composition des communautés, influencée par les conditions environnementales à petite échelle, la distance à la côte et la dynamique des courants. Près des îles principales, les communautés bénéficient de processus de fertilisation, avant qu'une partie d'entre elles ne soit transportée plus au large, initiant parfois un second « bloom ». L'effet d'île (EI) des Marquises apparaît ainsi comme un processus constant d'enrichissement, structurant spatialement les communautés phytoplanctoniques.
L'étude satellite à grande échelle, la modélisation ainsi que le suivi des flotteurs physique-biogéochimique (thèse de T. Hermilly) combinés à l'étude des macro- et micronutriments et des communautés phyto- et zooplanctoniques (stocks et flux de C et N), vont être essentielles pour aller plus loin dans l'étude de la structure et du fonctionnement de l'écosystème pélagique autour des Marquises et de l'IE.
Mesures acoustiques et distribution du micronecton (Barbin L., 2019 ; Barbin et al., 2020)
Les densités acoustiques acquises par enregistrement acoustique à 38kHz, permettent d'étudier l'abondance et répartition du micronecton ; Sv (densités par unité de volume) est un indicateur de la densité des organismes et le NASC (densités par unité de surface) est un indicateur de la biomasse des organismes. Les densités acoustiques (Sv) en été pendant MOANA-MATY 2019 montrent un fort gradient latitudinal entre Tahiti et les Marquises avec de fortes valeurs dans la zone proche des îles Marquises (zone HNLC) et des minimums dans la zone oligotrophe entre les Marquises et Tahiti (Fig. 13). Des migrations verticales du micronecton sont visibles sur l'ensemble des données avec la présence de deux couches diffusantes à fortes densités acoustiques ou DSL (Deep Scattering Layer) entre 0-200m et 400-600m, et un approfondissement de la DSL dans la zone oligotrophe. Ces résultats sont comparables à ceux observés en hiver austral (MOANA-MATY 2018) indiquant une certaine stabilité saisonnière à l'échelle régionale de la répartition du micronecton tant dans la différence entre la zone oligotrophe à HNLC que dans la profondeur des couches diffusantes.
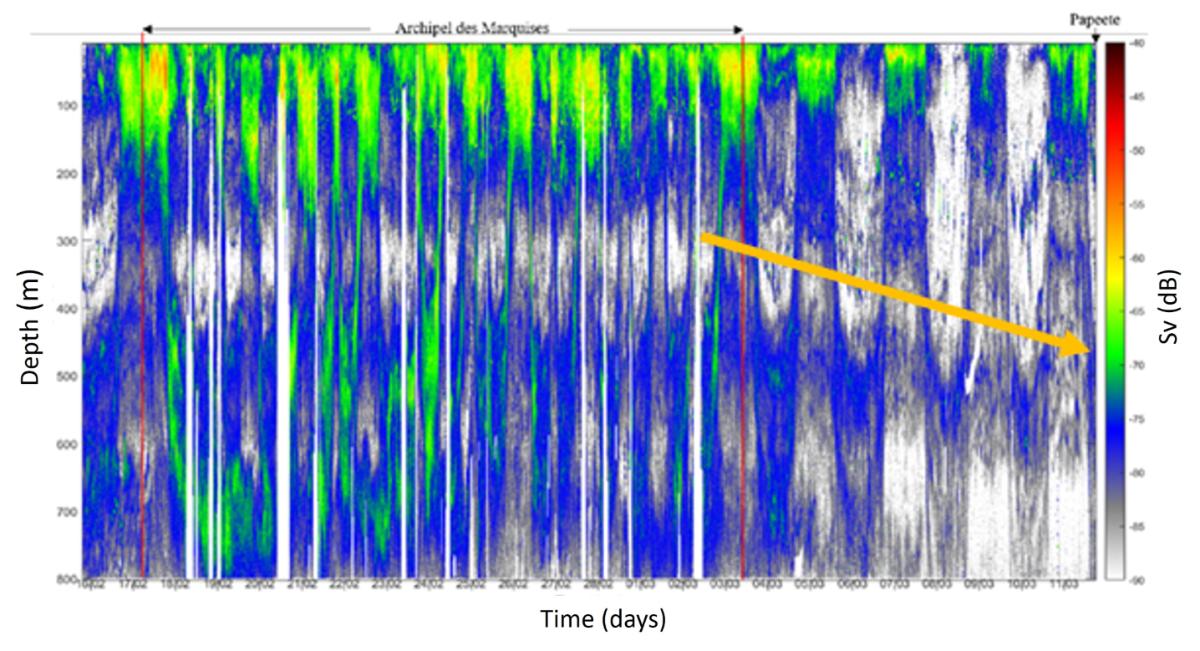
Figure 13. Densités acoustiques (Sv) en fonction du temps au cours de la campagne MOANA-MATY 2019, autour de l'archipel des Marquises et sur le trajet retour sur Tahiti.
Autour de l'archipel des Marquises, les densités acoustiques (NASC) (Fig. 14) sont en moyenne plus fortes sous le vent des îles (SO, aval), reflétant l'influence marqué du SEC, s'écoulant vers l'ouest, sur la répartition spatiale des organismes. Ces résultats contrastent avec ceux de 2018 en hiver, où les densités acoustiques étaient réparties équitablement autour de l'archipel en lien avec une courantologie plus "perturbée". Toutefois les données de courants de surface à elles seules ne suffisent pas à expliquer la distribution du micronecton autour de l'archipel, ni sa répartition verticale.

Figure 14. Profils verticaux de NASC moyens à 38kHz dans l'archipel, suivant un plan «amont/aval» (N/E vs S/O) des îles, défini par le trait rouge.
Les variables environnementales les plus importantes dans la structuration verticale du micronecton dans le Pacifique sud sont la concentration en oxygène dissous et la profondeur de la zone euphotique (Receveur et al., 2019). A l'échelle des Marquises, la comparaison entre les NASC et les paramètres environnementaux obtenus pendant MOANA-MATY 2019, mais également lors de la campagne de 2018, confirme que l'oxygène est le paramètre hydrologique qui impacte fortement la répartition verticale du micronecton. En effet, des minimums de NASC sont observés dans la zone de minium d'oxygène (300-400m), indiquant que les organismes évitent la zone hypoxique. A l'inverse, la salinité et la Chl-a semblent avoir une influence limitée et aucun lien clair n'a pu être établi entre la répartition du micronecton et la production primaire et la température au cours de cette étude.
Comprendre les liens entre la répartition du micronecton et les variables environnementales nécessitera d'améliorer les modéles physiques et biogéochimiques et d'utiliser des outils statistiques avancés de classification des profils CTD et acoustiques.
Liste des M1 et école d'ingénieurs ayant utilisé les données de MOANA-MATY 2 (M2 et thèse cf. section « Bibliographie)
- Joséphine Charavit (2019) Distribution spatiale de la Chlorophylle a et structure de taille du phytoplancton dans l'archipel des Marquises à deux saisons contrastées (campagnes océanographiques Moana-Maty 1 et 2. M1 Biologie Ecologie et Evolution, Aix-Marseille Université (encadrant M. Rodier - EIO)
- Jerome Castant (2020). Evaluation de la variabilité des masses d'eau dans l'archipel à partir des données des flotteurs Argo durant la période 2018-2019. M1 Université du littoral (encadrants C. Maes, E. Martinez - LOPS)
- Claire Le Roux (2020). Current climatology derived from Lagrangian drifters in the region of The Marquesas Islands. Ecole d'ingénieur Ecole centrale Lyon (encadrants C.Maes, G. Charria - LOPS)
- Claire Layec (2024). Influence of the island mass effect of the Marquesas on protist diversity. M1 Master Sciences de la mer et du littoral - Biologie, Communautés & Ecosystèmes, UBO (encadrant M. Sourisseau - IFREMER)
Références citées :
Barbin L., Habasque J., Martinez E., Rodier M. (2020). Acoustic data from MOANA MATY surveys. SEANOE. doi.org/10.17882/75317
Claustre, H., Sciandra, A., & Vaulot, D. (2008). Introduction to the special section bio-optical and biogeochemical conditions in the South East Pacific in late 2004: the BIOSOPE program. Biogeosciences, 5(3), 679-691.
Gove, J. M., McManus, M. A., Neuheimer, A. B., Polovina, J. J., Drazen, J. C., Smith, C. R., ... & Williams, G. J. (2016). Near-island biological hotspots in barren ocean basins. Nature communications, 7(1), 10581
Hasegawa, D., Lewis, M. R., & Gangopadhyay, A. (2009). How islands cause phytoplankton to bloom in their wakes. Geophysical Research Letters, 36, L20605. doi.org/10.1029/2009GL039743
Martinez, E., Rodier, M., & Maamaatuaiahutapu, K. (2016). Environnement océanique des Marquises. Biodiversité Terrestre et Marine des îles Marquises, Polynésie Française; Galzin, R., Duron, S.-D., Meyer, J.-Y., Eds, 123-136
Martinez, E., Rodier, M., Pagano, M., & Sauzede, R. (2020). Plankton spatial variability within the Marquesas archipelago, South Pacific. Journal of Marine Systems, 212, 103432.19
Rodier M. (2018) MOANA-MATY 2018 cruise, RV Alis, https://doi.org/10.17600/18000580
Signorini, S. R., McClain, C. R., & Dandonneau, Y. (1999). Mixing and phytoplankton bloom in the wake of the Marquesas Islands. Geophysical Research Letters, 26(20), 3121¿3124. doi.org/10.1029/1999GL010470
Torok, P., T-Krasznai, E., B-Beres, V., Bacsi, I., Borics, G., & Tothmeresz, B. (2016). Functional diversity supports the biomass-diversity humped-back relationship in phytoplankton assemblages. Functional Ecology, 30(9), 1593¿1602. doi.org/10.1111/1365-2435.12631
Receveur, A. (2019). Ecologie spatiale du micronecton: distribution, diversité et importance dans la structuration de l'écosystème pélagique du Pacifique sud-ouest (Doctoral dissertation, Aix-Marseille