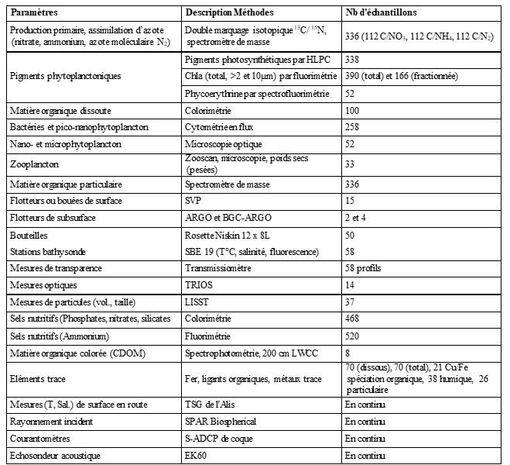
- Hydrodynamisme et dérive de particules: bouées SVPs versus courant de surface GEKCO (Cassianides et al., 2020; stages de Cassianides A., 2019; Le Roux C. 2020; Castant J., 2020)
15 bouées dérivantes de surface (SVP) ont été déployées pendant MOANA-MATY 1 (Fig.3), qui, en plus de fournir aux instances météorologiques des observations atmosphériques, ont permis d'évaluer la pertinence du produit de courants de surface GEKCO dérivés des observations satellites. Ces champs de courant sont la somme de la composante géostrophique (dérivée de l'élévation de la hauteur de surface de la mer) et de la composante d'Ekman (dérivée de la contrainte du vent).
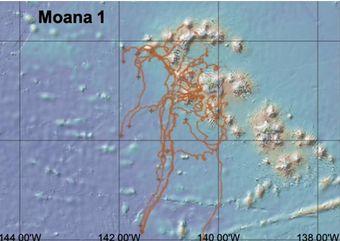
Fig.3. Trajectoires des bouées dérivantes de surface 15 jours après la mission
Des simulations lagrangiennes ont été effectuées grâce au logiciel ARIANE (Blanke et al., 1997), en lâchant tous les jours du 1er au 10 octobre 2018 des particules fictives dans les champs de courant GECKO au nord de l'archipel où ont été déployées les SVP (Fig.4). Leurs trajectoires ont été calculées pendant 3 mois puis comparées avec celles des SVP déployés pendant la mission. La dispersion des particules fictives simulée est plus cohérente avec celle des SVP lorsque la composante liée au vent est divisée par 2, indiquant une surestimation de la composante d'Ekman dans GEKCO et suggérant d'utiliser un produit corrigé de 50% de cette composante.
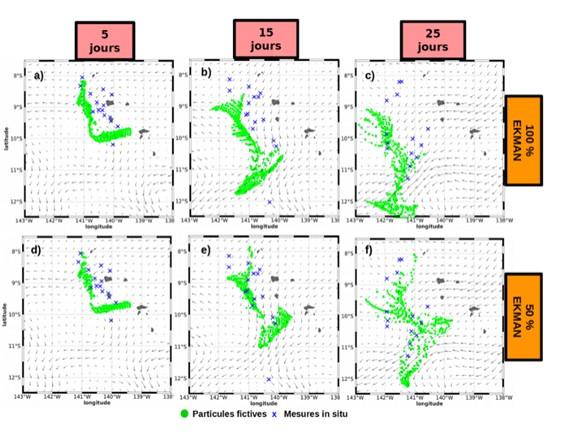
Fig. 4. Comparaison de la dispersion des bouées dérivantes de surface déployées lors de MOANA-MATY 1 (croix bleues) avec celle de particules fictives (cercles verts).
En complément des données obtenues à l'aide des bouées dérivantes, des mesures obtenues à partir du S-ADCP fixé à la coque du NO Alis (Ship borne Acoustic Doppler Current Profiler) ont permis d'établir une cartographie des courants au cours de la campagne (Fig. 5), montrant une forte activité dynamique autour des îles.
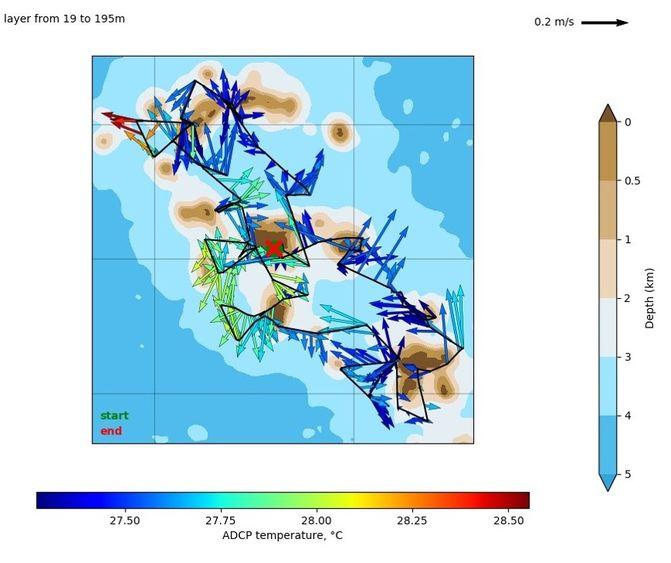
Fig. 5. Courants moyens sur la couche 19-195m enregistrés au cours de la campagne MOANA-MATY1
- Distribution du Fer autour des Marquises (Grand M. - Moss Landing Marine Labs/USA, atelier déc.2022)
La biodisponibilité du fer est un facteur limitant de la production phytoplanctonique dans de nombreuses régions océaniques. Un des objectifs de la campagne MOANA-MATY 1 était de mesurer cette biodisponibilité dans la zone des Marquises et tester l'hypothèse d'une limitation en fer dans cette région riche en macro-nutriments (NOx + NH4 = 4,21 ± 1,3 µmol L-1; PO4 = 0,48 ± 0,07 µmol L-1; Si(OH)4 = 2,0 ± 0,3 µmol L-1; valeurs moyennes pendant la campagne) . Des mesures de fer ont donc été réalisées sur 13 stations, ciblant cinq profondeurs entre 0 et 90m, à l'aide d'une pompe à soufflet en Téflon et d'un tuyau en Téflon nettoyé à l'acide. Les échantillons ont été analysés pour le Fer dissous (dFe: échantillons filtrés) et le Fer dissolvable (TDFe: échantillons non filtrés) au Moss Landing Marine Labs. La méthode d'analyse utilisée par chemiluminescence (modifié d'après Obata et al., 1993) a montré une limite de détection à 0,02 nM, avec une précision analytique de < 5% pour les [dFe] > 0,15 nM et de 5-15% pour des [dFe] < 0,15 nM, et des taux de récupération > 95% pour D1 (n=18) et GSC (n=4) par rapport aux échantillons de référence SAFe et GEOTRACES.
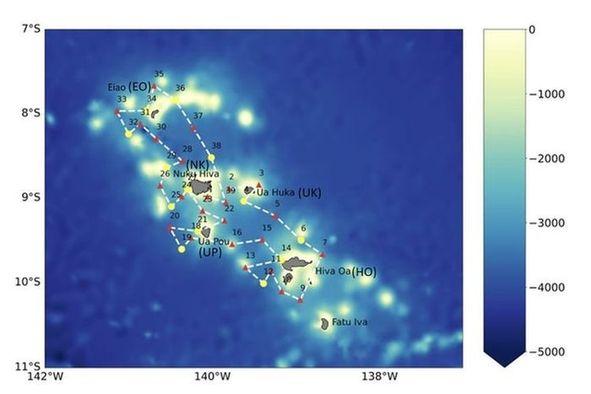
Les résultats ont révélé que la concentration moyenne de dFe en surface pendant MOANA-MATY 1 était de 0,12 ± 0,05 nM, en accord avec des études antérieures telles que Blain et al. (2008). Bien que la majeure partie de la zone d'étude ait montré des [dFe] en surface uniformément basses (~ 0,15 nM), deux stations spécifiques, une située au nord-ouest de Nuku Hiva (triangle orange) et une sur le côté ouest d'Ua Pou (croix verte), ont affiché des concentrations élevées allant jusqu'à 0,25 nM sur l'ensemble de la colonne 0-90m (Fig. 6).
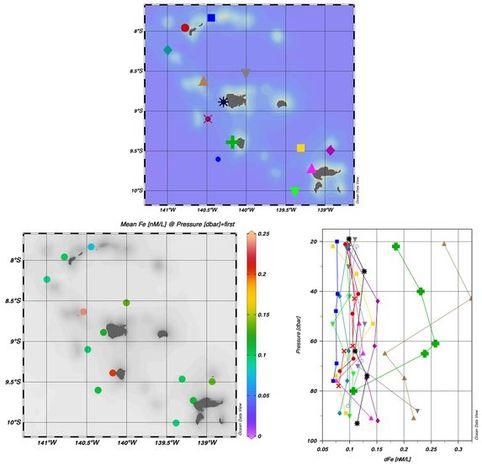
Fig. 6. Fer dissous (dFe) mesuré au cours de MOANA-MATY 1. Bas gauche: moyennes en surface, Bas droit: profiles verticaux
Les données TDFe, englobant à la fois le dFe et le Fer labile associé aux particules biogéniques et lithogéniques, ont mis en évidence des valeurs accrues sur les périphéries ouest de toutes les îles (Fig.7). Cela suggère une contribution potentielle, bien que localisée, de Fer provenant des sédiments des îles. Des échantillons d'eau prélevés dans des ruisseaux à Eiao et Ua Pou ont par ailleurs montré des concentrations de Fer de 82 nM et 280 nM, respectivement. La variance des concentrations pourrait être attribuée à des lithologies différentes entre ces deux îles. En collaboration avec Helene Planquette (LEMAR), ces informations seront intégrées aux données de métaux traces particulaire pour élucider l'influence potentielle des sédiments insulaires dans la fourniture de Fer biodisponible (travail en cours).
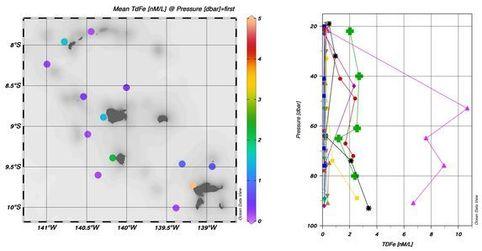
Fig. 7. Fer total dissolvable (TDFe) pendant MOANA-MATY 1. Bas gauche: moyennes en surface, Bas droit: profiles verticaux.
- Activité hydrodynamique et distribution du phytoplancton : données satellites et in situ (Cassianides et al., 2020; Thèse de Hermilly T. 2022-2025; stages de Charavit J., 2019 ;Matitia T., 2019)
Deux efflorescences phytoplanctoniques remarquables ont été observées à partir des images satellite GlobColor-GSM, pendant et juste après la campagne et ont été étudiées en combinant les observations satellites et données in-situ issues de la campagne MOANA-MATY 1. A titre d'exemple (Fig. 8), les observations de courants de surface combinées à une approche lagrangienne illustrent le fait que les particules observées au niveau du panache de Chla le 3 Octobre 2018 au sud de l'archipel sont passées à proximité directe des îles (95% des cas), indiquant que les îles sont à l'origine même des enrichissements biologiques observés. La circulation méso-échelle modifie ensuite la distribution du panache de Chla sur plusieurs centaines de kilomètres en aval des îles.
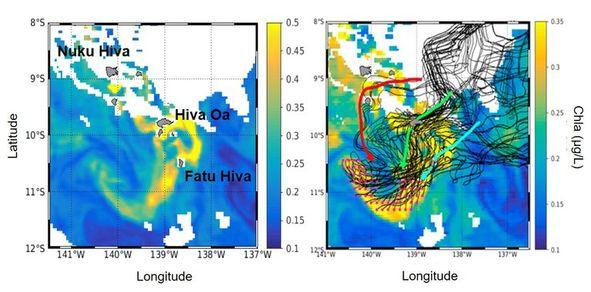
Fig. 8. Gauche : Cartes de Chla (µg L-1) dérivées des images de couleur de l'océan du produit GlobColor-GSM le 3 oct. 2018. Droite : Chla et trajectoires de dérive des particules fictives (trait noir) issue d'une analyse lagrangienne en mode backward depuis leurs positions initiales (points magenta). Les trois flèches indiquent les chemins préférentiels empruntés par les particules, autour de Ua Huka, Nuku Hiva et Ua Pou (rouge), Hiva Oa (vert), Fatu Hiva (bleu).
En accord avec les données satellites, la distribution de la Chla in situ (Fig. 9) montre que les plus forts enrichissements ont lieu près des îles dans la couche de surface (0,80 à 0, 90 µg Chla L-1) et de préférence sous le vent des îles, indiquant des conditions environnementales favorables au développement du phytoplancton (apports de macro- et micronutriments, stabilité de la colonne d'eau, etc ...). Toutefois, due à la forte activité dynamique à mésoéchelle observée en hiver austral pouvant modifier les panaches (Fig. 8), le contraste entre les valeurs de Chla au vent et sous le vent des îles est peu marqué. En dehors des zones enrichies, la Chla est distribuée plus en profondeur dans la couche euphotique avec un maximum (DCM) entre 60-100m et les valeurs de production restent faibles.
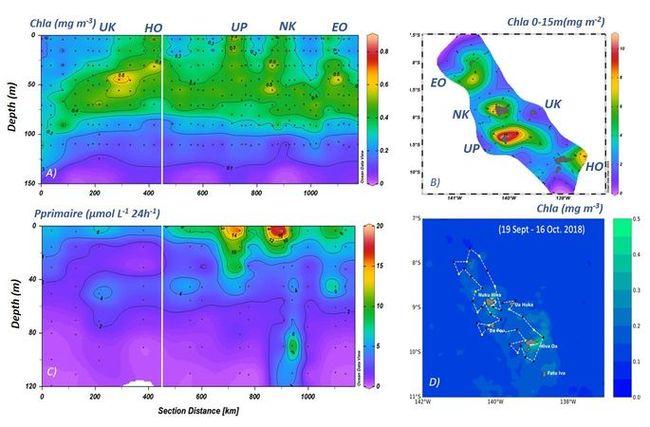
Fig. 9. Distribution de la Chla et production primaire en hiver austral au cours de la campagne MOANA-MATY 1. A) Chla total par HPLC, B) Chla integrée sur 0-15m C) Primary production D) Chla de surface dérivée des images satellite GlobColor-GSM. Le trait blanc sur A et C indique la zone au vent des îles (à gauche) et sous le vent des îles (droite).
Les valeurs intégrées de Chla sur la couche euphotique (0,1% de lumière) varient entre 10,4 et 52,2 mg Chla m-2 (30,8 ± 9,2 mg Chla m-2). Elles sont associées à des valeurs de production primaire variant de 144 à 682 mgC m-2 j-1 (320 ± 184 mgC m-2 j-1) et maximales en surface (0-20/30m) et près de Ua Pou (Fig. 9). Ces valeurs traduisent un système mésotrophe (de type HNLC) si on les compare aux données de l'upwelling équatorial entre 140°W et 110°W de l'ordre de 26,5 ± 3,5 mg Chla m-2 et 748 ± 110 mg C m-2 j-1 (Balch et al., 2011). L'écosystème pélagique autour de l'archipel des Marquises apparait donc comme un système globalement productif mais sujet à une forte variabilité à plus petite échelle de temps et d'espace, liée en grande partie à la forte et complexe activité dynamique océanique.
La biomasse chlorophyllienne est largement dominée par le picoplancton (Chla < 2µm représentant 75 à 95% de la Chla totale), comme dans l'upwelling équatorial (Blach et al., 2011), et en accord avec les observations antérieures (Martinez et al. 2012). Les données pigmentaires (HPLC) montrent une prédominance des picocyanobactéries (40 à 60% de la Chla totale), largement dominées par les Synechococcus en surface près des îles avec des abondances pouvant dépasser 80 ×103 cellules mL-1, alors que les Prochloroccoccus dominent par ailleurs (données de cytométrie en flux). Les observations en microscopie inversée ont également permis de dresser un premier inventaire taxonomique du nano- et microphytoplancton ainsi que de mettre en évidence des changements de communautés au sein de l'archipel avec des maxima d'abondances à proximité des îles et près de Ua Pou (Fig. 10). Les flagellés dominent avec deux principaux groupes, les Prymnésiophytes (16 espèces mais principalement des Coccolithophores) et les Dinoflagellés (81 espèces). Les diatomées (65 espèces) sont plus abondantes en aval des îles avec un maximum près de Ua Pou. Ces observations microscopiques, pigmentaires et en cytométrie en flux seront complétées/confirmées par des mesures d'ADN meta-barcoding lors de la seconde mission MOANA-MATY 2.
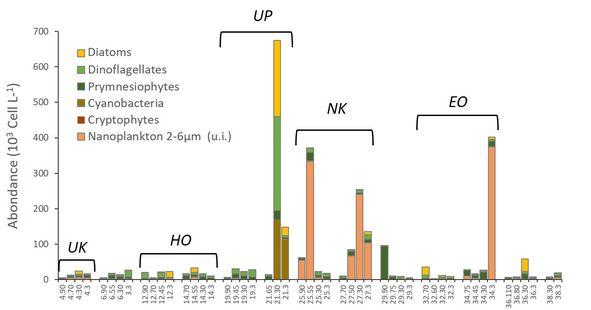
Fig. 10. Composition des communautés nano-et microphytoplanctoniques en hiver austral pendant MOANA-MATY 1 (observations au microscope inversé). En abscisse #station-profondeur (en m). Ua Huka (UK) , Hiva Oa (HO), Ua Pou (UP), Nukku Hiva (NK), Eiao (EO).
L'ensemble des données phytoplanctoniques pose les questions de savoir 1) quelle est la part de Chla observée dans les panaches qui est directement advectée depuis les îles et celle potentiellement induite sur place par la dynamique propre aux tourbillons ou aux zones de fronts.et, 2) comment la structure des communautés phytoplanctonique va structurer l'ensemble du réseau trophique. L'étude satellite à grande échelle, la modélisation ainsi que le suivi des flotteurs Argo physique-biogéochimique (thèse de T. Hermilly, 2022-2025) combinés à l'étude des macro- et micronutriments et des communautés phyto- et zooplanctoniques (stocks et flux de C et N), vont être essentielles pour comprendre l'origine des enrichissements et aller plus loin dans l'étude du fonctionnement de l'écosystème pélagique autour des Marquises.
- Mesures acoustiques et distribution du micronecton (Barbin et al., 2020; stage de Barbin L., 2020)
Les densités acoustiques acquises par enregistrement acoustique à 38kHz, permettent d'étudier l'abondance et la répartition du micronecton. Ces données montrent un fort gradient latitudinale entre Tahiti et les Marquises avec de fortes valeurs dans la zone proche des îles Marquises et des minima dans la zone oligotrophe lors des transits aller et retour depuis/vers Tahiti (Fig. 11). Les migrations verticales des organismes micronectoniques sont visibles sur l'ensemble des données avec la présence de deux couches diffusantes à fortes densités acoustiques (Deep Scattering Layer DSL) entre 0-200m et 400-600m.
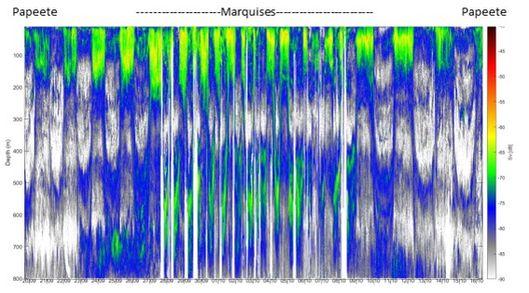
Fig. 11. Intensité acoustique par unité de volume (volume backscattering coefficient, Sv en dB) sur la colonne d'eau en fonction du temps pendant MOANA-MATY 1. Les pointillés délimitent la partie de la campagne dans l'archipel des Marquises
Au niveau des Marquises, les données acoustiques par unité de surface (NASC, nautical area scattering coefficient, Fig. 12) sont réparties équitablement autour de l'archipel de jour comme de nuit, n'indiquant pas de différences significatives entre les zones au vent («amont ») et sous le vent (« aval »).
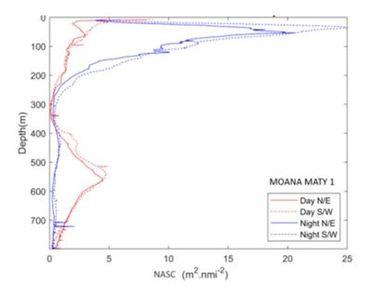
Fig. 12. Profils verticaux de NASC moyens à 38kHz dans l'archipel, suivant un plan «amont/aval» des îles (N/E vs S/W).
Les variables environnementales les plus importantes dans la structuration verticale du micronecton dans le Pacifique sud décrites par Receveur et al. (2019) sont la quantité d'oxygène dissous et la profondeur de la zone euphotique. La comparaison entre les NASC et les paramètres environnementaux à l'échelle de l'archipel semble indiquer que l'oxygène est le paramètre hydrologique qui impacte le plus la répartition verticale du micronecton. A l'inverse, la dynamique océanique dans l'archipel semble jouer un rôle mineur. Les résultats soulignent toutefois la complexité de la répartition du micronecton autour des îles et la difficulté à établir des liens directs entre sa répartition et les paramètres environnementaux.
Liste des stages de M1 et école d'ingénieurs ayant utilisé les données de MOANA-MATY 1 (M2 et thèse cf. "Références")
Charavit Joséphine (2019) Distribution spatiale de la Chlorophylle a et structure de taille du phytoplancton dans l'archipel des Marquises à deux saisons contrastées (campagnes océanographiques Moana-Maty 1 et 2. M1 Biologie Ecologie et Evolution, Aix-Marseille Université (encadrante M. Rodier - EIO)
Matitia Tanya (2019). Effet d'iles aux Marquises : étude de la variabilité spatiale et saisonnière des paramètres physiques et biogéochimiques dans l'archipel. 3rd yr of Bachelor, Université Montréal, Canada (encadrantes M. Rodier - EIO & E. Martinez - LOPS)
Castant Jerome (2020). Evaluation de la variabilité des masses d'eau dans l'archipel à partir des données des flotteurs Argo durant la période 2018-2019. M1 Université du littoral (encadrants C. Maes & E. Martinez - LOPS)
Le Roux Claire (2020). Current climatology derived from Lagrangian drifters in the region of The Marquesas Islands. Ecole d'ingénieur Ecole centrale Lyon (encadrants C.Maes & G. Charria - LOPS)
Références citées
Balch, W. M., Poulton, A. J., Drapeau, D. T., Bowler, B. C., Windecker, L. A., & Booth, E. S. (2011). Zonal and meridional patterns of phytoplankton biomass and carbon fixation in the Equatorial Pacific Ocean, between 110 W and 140 W. Deep Sea Research Part II: Topical Studies in Oceanography, 58(3-4), 400-416
Blain, S.; Bonnet, S.; Guieu, C. (2008) Dissolved iron distribution in the tropical and sub tropical South Eastern Pacific. Biogeosciences, 5, 269¿280.
Blanke, B.; Raynaud, S. (1997) Kinematics of the Pacific Equatorial Undercurrent: An Eulerian and Lagrangian Approach from GCM Results. Journal of Physical Oceanography, 27(6), 1038-1053.
Claustre, H.; Sciandra, A.; Vaulot, D. (2008) Introduction to the special section bio-optical and biogeochemical conditions in the South East Pacific in late 2004: The BIOSOPE program. Biogeosciences, 5, 679¿691.
Martinez E., Rodier M., Maamaatuaiahutapu K. (2016) Environnement océanique des Marquises. In Galzin R., Duron S.-D. & Meyer J.-Y. (eds), Biodiversité terrestre et marine des îles Marquises, Polynésie française. Société Française d'Ichtyologie, Paris : 123-136.
Martinez, E., Raapoto, H., Maes, C., Maamaatuaihutapu, K. (2018) Influence of Tropical Instability Waves on Phytoplankton Biomass near the Marquesas Islands. Remote Sensing, 10(4), 640.
Martinez E., Rodier M., Pagano, M., Sauzède R. (2020) Plankton spatial variability within the Marquesas archipelago, South Pacific. Journal of Marine Systems, 212, 103432
Obata, H., Karatani, H., Nakayama, E. (1993). Automated determination of iron in seawater by chelating resin concentration and chemiluminescence detection. Analytical Chemistry, 65(11), 1524-1528.
Receveur, A. (2019). Ecologie spatiale du micronecton: distribution, diversité et importance dans la structuration de l'écosystème pélagique du Pacifique sud-ouest (Doctoral dissertation, Aix-Marseille).