Le principal objectif scientifique de MOBYDICK est la compréhension de la pompe biologique de carbone océanique en (i) considérant les réseaux alimentaires marins de manière « end to end », et (ii) en reliant la biodiversité aux flux biogéochimiques. Pour atteindre ces objectifs, nous avons étudié des types d'écosystèmes contrastés dans l'océan Austral : un écosystème à faible biomasse et faible exportation et un écosystème à forte biomasse et faible exportation dans la région de l'île Kerguelen (Fig. 1). La biomasse plus élevée à l'est de l'île de Kerguelen est due à la fertilisation naturelle en fer dans des eaux à forte teneur en nutriments et à faible teneur en chlorophylle (HNLC). Les observations in situ et les analyses des échantillons collectés au cours de la campagne ont permis de décrire en détail les structures des communautés aux différents niveaux trophiques dans des réseaux trophiques marins contrastés. Par rapport aux études précédentes, l'originalité de MOBYDICK est l'étude simultanée de la biodiversité pélagique à tous les niveaux trophiques et de presque tous les groupes fonctionnels, des bactéries (bactéries et archées) aux prédateurs supérieurs, ainsi que l'acquisition de données sur les facteurs environnementaux, notamment les nutriments dissous et particulaires.
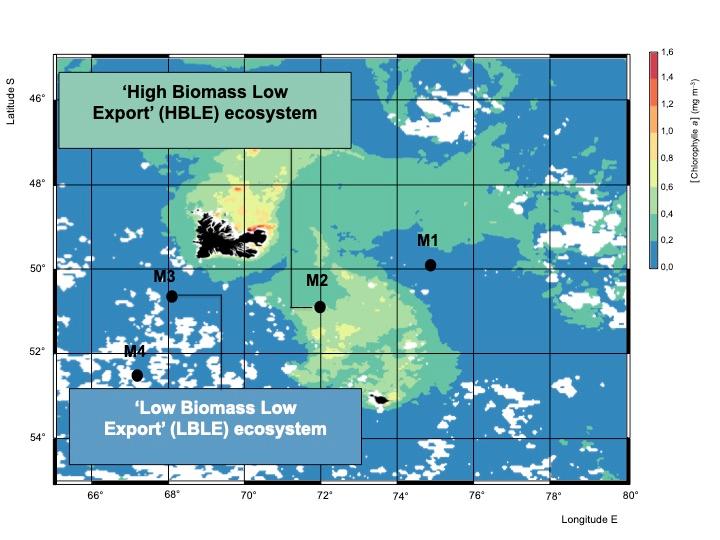
Fig. 1. Sites d'étude dans des écosystèmes contrastés, définis par leurs régimes de productivité.
Dans ce qui suit, nous décrirons brièvement un résultat majeur obtenu pour chacun des niveaux trophiques considérés et fournirons une vue intégrée des flux biogéochimiques associés sur les sites contrastés.
Une mosaïque de fonctions bactériennes spécifiques à la taxonomie dans l'utilisation du Fe et du C
Un litre d'eau de mer contient environ 1012 gènes bactériens (incluant les bactéries et les archées), ce qui illustre l'immense potentiel métabolique présent dans les microorganismes marins. Mais la façon dont les fonctions associées sont réparties entre les taxons bactériens, et comment la diversité bactérienne affecte le cycle des éléments, reste une question mal comprise. La campagne MOBYDICK a fourni une excellente occasion de collecter des échantillons pour une étude complète et approfondie de la structure et de la fonction de la communauté bactérienne, en utilisant une gamme de technologies -omiques modernes (métagénomique, -transcriptomique et -protéomique). Dans ce contexte, une approche relativement récente permet de reconstruire des taxons bactériens individuels à partir de séquences métagénomiques environnementales. Ces "génomes assemblés du métagénome" (MAG) fournissent des informations sur le répertoire et l'expression des gènes au niveau de l'"espèce" bactérienne, informations qui étaient jusqu'à présent limitées aux souches cultivées. La reconstruction de 133 MAG à partir des séquences MOBYDICK nous a permis de décrire les stratégies très diverses des différents taxons bactériens en considérant les voies métaboliques centrales du métabolisme du Fe et du C (Fig. 2)(Sun et al. 2021). Des MAGs taxonomiquement proches pourraient présenter des propriétés contrastées de régulation de l'expression des gènes ou une distribution distincte de l'abondance des gènes entre les sites. Cela indique une adaptation et une spécialisation à des niches écologiques différentes. Nos résultats fournissent une image détaillée des rôles fonctionnels des taxons bactériens individuels dans les environnements contrastés de l'océan Austral et ils soulignent l'interaction complexe entre le répertoire génétique des taxons individuels et leur environnement.
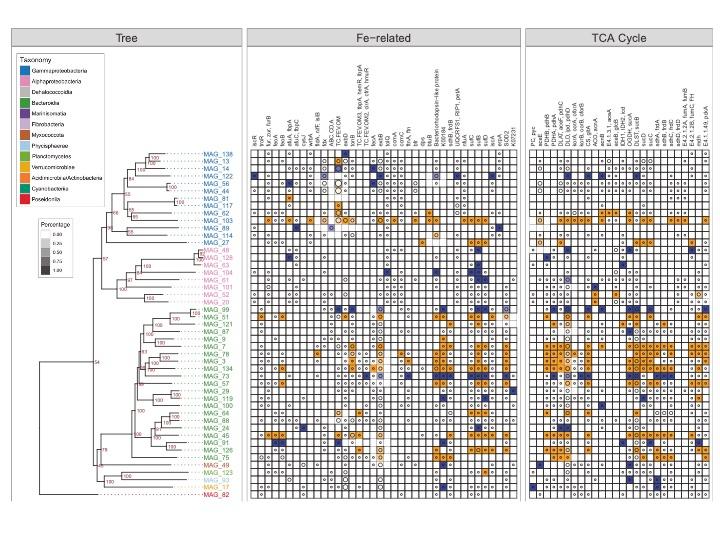
Fig. 2. Illustration de la présence, de l'abondance et de l'expression des gènes dans les génomes assemblés métagénomiques (MAGs) obtenus au cours de la campagne MOBYDICK. De gauche à droite, les panneaux représentent l'arbre phylogénétique, les gènes liés à l'absorption bactérienne de Fe et les gènes impliqués dans le cycle de l'acide tricarboxylique (TCA), une voie centrale du métabolisme cellulaire du carbone. Chaque bloc carré décrit les statistiques d'une famille de protéines dans un MAG. Un carré vide suggère qu'aucun gène du MAG (axe y) n'est classé dans le groupe fonctionnel correspondant (axe x). Un cercle dans le bloc carré indique l'identification d'homologues d'une famille de protéines dans le MAG, sa taille étant proportionnelle au nombre de gènes affectés à cette famille. Les blocs carrés sont colorés en fonction des profils d'expression différentielle de leur(s) gène(s). Les gènes, dont l'expression est significativement plus élevée dans le site M2 fertilisé par le Fe par rapport aux sites M3 et M4 HNLC, sont mis en évidence en orange ; vice versa, en bleu. D'après Sun et al. (2021).
Fixation du CO2 par le petit phytoplancton : aperçu des observations unicellulaires
Le phytoplancton est composé de micro-organismes caractérisés par une large gamme de tailles et une grande diversité phylogénétique. La contribution de chaque groupe phytoplanctonique à la fixation du CO2 dans les eaux de surface et à l'exportation du carbone en profondeur n'est pas bien connue. La détermination de la variabilité intra- et inter-groupe de la fixation du CO2 est cruciale pour comprendre le rôle des différents groupes phytoplanctoniques dans la pompe biologique du carbone. Au cours de la campagne MOBYDICK, nous avons mesuré les taux de fixation du CO2 de différents groupes de phytoplancton au niveau de la cellule unique, en utilisant des isotopes stables (13CO2) et la spectrométrie de masse des ions secondaires à l'échelle nanométrique (NanoSIMS) (Irion et al. 2021)(Fig. 3). Nous avons observé qu'à la fin de l'été austral, lorsque la campagne MOBYDICK a eu lieu, les petites cellules (< 20 µm) composées de taxons phylogénétiquement éloignés (prymnésiophytes, prasinophytes et petites diatomées) se développaient plus rapidement que les diatomées plus grandes. Ces dernières présentaient une croissance hétérogène et une proportion considérable de cellules inactives (19±13%). Par conséquent, les petits phytoplanctons ont largement contribué à la fixation totale du CO2 (41-70%). La distribution verticale des pigments indique que la prédation pourrait être une voie d'exportation importante pour le carbone fixé par le petit phytoplancton. Ensemble, nos résultats soulignent le rôle important des petites cellules phytoplanctoniques dans la fixation du CO2 dans l'océan Austral. Ces résultats complètent l'image saisonnière dans la région naturellement fertilisée en fer de l'île de Kerguelen, où les diatomées ont été identifiées comme des acteurs majeurs dans l'absorption et l'exportation de CO2 au printemps et en été.
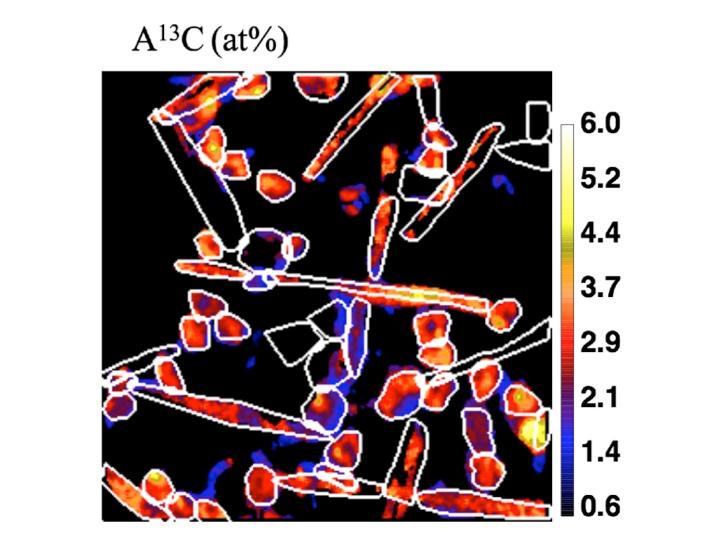
Fig. 3. Analyse de l'échantillon par NanoSIMS (Nanoscale secondary ion mass spectrometry). Le rapport 13C/12C, indiqué par le code couleur (axe des ordonnées) est utilisé pour quantifier la fixation du CO2 des cellules individuelles triées par cytométrie de flux. D'après Irion Solène (thèse de doctorat).
Les diatomées en route vers l'océan profond
Malgré l'importance de l'exportation de carbone organique vers les profondeurs via les particules, la composition des divers types et gammes de tailles de particules présentes dans les différentes couches profondes est encore largement inconnue. Un défi méthodologique est l'utilisation d'instruments appropriés pour collecter les particules afin de les caractériser et de les quantifier. Au cours de MOBYDICK, nous avons déployé un dispositif récemment disponible qui concentre et collecte les particules en profondeur sur une couche d'eau prédéterminée, un échantillonneur appelé BottleNet. Ce premier déploiement de l'échantillonneur BottleNet a permis de fournir une description détaillée des particules et des cellules planctoniques individuelles, y compris leur taxonomie, leur contenu en carbone et en lipides, ainsi que leur viabilité dans différentes couches de profondeur (par exemple 60-125m, 125-500m et 500-1500m) (Leblanc et al. 2021). De manière inattendue, la majorité des particules collectées étaient constituées de frustules de diatomées vides, alors que les pelottes fécales et les agrégats ne représentaient qu'une fraction mineure (Fig. 4). Nous avons identifié l'occurrence de processus de mortalité distincts pour les taxons de diatomées, allant de l'infection parasitaire au broutage par le mésozooplancton, ainsi que des degrés de silicification et des stades de vie distincts, et avons ainsi pu mettre en évidence des modes d'exportation distincts vers les couches intermédiaires et profondes.

Fig. 4. Pelote fécale contenant des débris reconnaissables de la diatomée Fragilariopsis kerguelensis (rose), et des diatomées centriques (turquoise). Quelques coccolithes sont également visibles (violet). D'après Leblanc (2021).
Dynamique saisonnière du réseau trophique microbien
La combinaison des observations réalisées au cours de la campagne MOBYDICK à la fin de l'été avec celles des projets précédents réalisés au début du printemps (KEOPS2) et en été (KEOPS1) nous a permis de dessiner un scénario saisonnier pour les ?ux du réseau trophique microbien dans les eaux fertilisées par le fer et les eaux HNLC (Christaki et al. 2021)(Fig. 5). Au début du printemps, le système est très productif, avec une faible ef?cience d'exportation de carbone, ce qui se traduit par une grande quantité de carbone phytoplanctonique potentiellement disponible pour les niveaux trophiques supérieurs. Pendant le déclin du bloom, la production brute de la communauté diminue de 3 fois par rapport au début du printemps. Le système est caractérisé par une exportation de carbone et une efficacité d'exportation de carbone élevées, tandis que le carbone phytoplanctonique potentiellement disponible pour les niveaux trophiques supérieurs est 10 fois plus faible par rapport au printemps. Pendant la phase post-floraison (MOBYDICK), la production brute de la communauté est similaire à celle déterminée pendant la phase de floraison déclinante. Cependant, l'exportation de carbone et l'efficacité de l'exportation sont à nouveau faibles, ce qui entraîne une biomasse disponible pour les niveaux trophiques supérieurs environ 2 fois plus élevée par rapport au déclin du bloom. Le rôle du réseau trophique microbien pour la transformation du carbone varie au cours des trois phases. Au début du printemps, une fraction modérée de la production primaire est canalisée par les bactéries (21%) et en raison d'une faible efficacité de croissance bactérienne (9%), cette matière organique est principalement respirée. Pendant le déclin du bloom, les bactéries hétérotrophes traitent une fraction plus élevée de la production primaire (44%) avec une production de biomasse bactérienne plus efficace par rapport au printemps. L'importance de la lyse virale dans les pertes de carbone entraîne un transfert inefficace de la biomasse bactérienne vers les nanoflagellés hétérotrophes. Pendant la phase post-efflorescence, le transfert de la biomasse bactérienne vers les nanoflagellés hétérotrophes (HNF) domine sur la perte par lyse virale. Prises ensemble, ces observations suggèrent un réseau trophique microbien globalement moins efficace au début du printemps et de l'été, la respiration et la lyse virale, respectivement, représentant des termes de perte importants. Pendant la période post-efflorescence, environ 30 % de la production brute de la communauté est transféré de manière efficace à la biomasse bactérienne et aux HNF, et sont donc potentiellement disponibles pour les niveaux trophiques supérieurs.
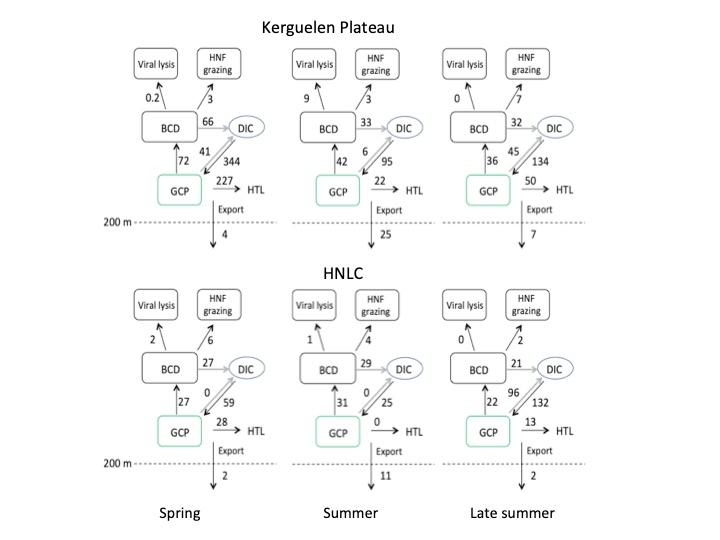
Fig. 5. Flux de carbone intégrés sur la couche de mélange au sein du réseau trophique microbien. De gauche à droite : au début (KEOPS2), à la fin (KEOPS1) et après l'efflorescence de diatomées (MOBYDICK). Une description détaillée des calculs est fournie dans la publication de Christaki et al. (2021).
Diversité et contribution au cycle du carbone des métazoaires
Alors que l'effet de la fertilisation naturelle en fer sur les cycles biogéochimiques et la structure du réseau trophique de niveau inférieur a été intensivement étudié au cours de la dernière décennie dans la région de Kerguelen, les niveaux trophiques supérieurs n'ont pas encore été inclus pour une approche complète de l'écosystème. En utilisant l'identification taxonomique macro- et micro-scopique des organismes collectés avec des filets et des chaluts, et des mesures acoustiques (18 et 38 kHz), nous avons pu décrire la structure et la fonction des réseaux alimentaires pélagiques jusqu'au micronecton. Les communautés de macrozooplancton et de micronecton différaient selon les régimes de productivité distincts, malgré un système vertical à trois couches bien visible, détectable dans les 800 m supérieurs sur tous les sites, à savoir les couches de diffusion à faible profondeur (10 à 200 m), à moyenne profondeur (200 à 500 m) et à grande profondeur (500 à 800 m) (Cotté et al. 2021)(Fig. 6). Alors que les salpes (Salpa thompsoni) dominaient la biomasse sur le plateau productif de Kerguelen, elles étaient rares dans les conditions HNLC. En outre, les crustacés (principalement Euphausia vallentini et Themisto gaudichaudii) étaient particulièrement abondants sur le plateau, représentant un stock de carbone important, bien que variable, dans la couche d'eau de 0-500 m. Les poissons mésopélagiques étaient proéminents sur le plateau. Les poissons mésopélagiques étaient proéminents en dessous de 500m et formaient des couches permanentes ou migrantes représentant la principale source de carbone. Nos observations à haute fréquence ont permis d'identifier des modèles spatiaux et temporels dans les distributions verticales du micronecton et le contenu en carbone associé. Nous avons également estimé les flux de carbone respirés par les myctophidés migrateurs, ce qui a permis de comprendre les principaux composants et mécanismes de l'exportation active de carbone dans la région et la façon dont ils sont modulés par la topographie complexe et les effets de la masse terrestre.
Les isotopes stables utilisés (delta15N, delta13C) combinés aux identifications taxonomiques ont révélé que la longueur de la chaîne alimentaire, calculée à partir d'un sous-ensemble standardisé d'espèces et de classes de taille, était plus courte d'un niveau trophique dans les eaux du plateau de Kerguelen fertilisées en fer que dans les eaux HNLC. Les rapports de masse prédateur-proie (PPMR) étaient inférieurs d'un ordre de grandeur dans la région HNLC (PPMR = 170) par rapport au plateau (PPMR > 2000), ce qui indique un recyclage interne élevé dans les eaux HNLC plus stables. L'efficacité globale du transfert du carbone et de l'azote a été estimée comme étant environ 2 fois plus élevée (21 %) dans la région HNLC que dans l'écosystème soutenu par la fertilisation naturelle en fer (7-10 %) (Hunt et al. 2021). Il est intéressant de noter que ces observations des niveaux trophiques supérieurs sont en accord avec celles des réseaux trophiques microbiens (Christaki et al. 2021), illustrant un fonctionnement différent de l'ensemble du réseau alimentaire dans les écosystèmes à forte et faible productivité de l'océan Austral.
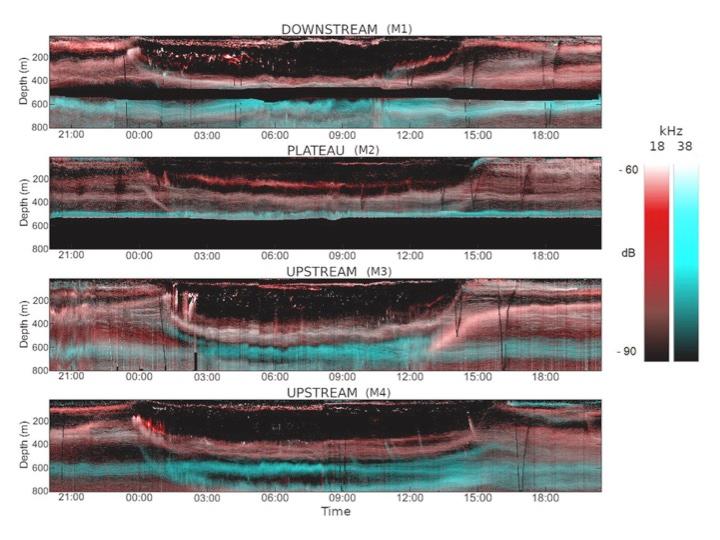
Fig. 6. Acoustique - composites quotidiens Rouge Vert Bleu (RGB) des intensités de rétrodiffusion volumique (dB re m-1) de 12 à 800m pour les quatre stations MOBYDICK (D'après Cotté et al. 2021).
