Grâce à un effort d'échantillonnage conséquent multispécifique, nous avons pu montrer que la diversité benthique des iles Loyautés était d'une grande richesse. Pour de nombres espèces, les échantillons collectés lors de la campagne BIBELOT sont venus compléter ceux de la mission COBELO (2013-ALIS), et ceux d'une mission précédente réalisée sur le bateau de la Living Ocean Fundation (en 2013). Ces échantillons ont ensuite été complétés par ceux de la campagne CHEST (2015-ALIS). Cet échantillonnage a permis d'estimer chez certaines espèces (principalement bénitiers, hydraires, et coraux du genre Pocillopora) les échelles spatiales de dispersion larvaire et de connectivité des populations.
En ce qui concerne les résultats majeurs obtenus à partir des échantillons récoltés lors de la mission BIBELOT et complétés par les autres échantillons des missions précédemment citées, on note des résultats remarquables pour tous les groupes benthiques sur lesquels nous nous sommes focalisés.
1) Les bénitiers
Pour les bénitiers, nous avons mis en évidence (grâce aux analyses génétiques réalisées sur les biopsies récoltées lors de la mission) la présence d'une nouvelle espèce de bénitier Tridacna noae (Figure 1), qui avait été récemment décrite endémique de Taiwan (Su et al. 2014), et que nous avons trouvé dans les Loyautés et le Nord de la Grande Terre, alors qu'elle semble absente du reste du territoire (Borsa et al. 2015a, Borsa et al. 2015b). Plus d'informations relatives à cette espèce sont disponibles dans l'article publié dans la lettre d'information sur les pêches de la CPS n° 145 et disponible, en français (Wabnitz & Fauvelot 2014)

Figure 1 : Photographie sous-marine de l'espèce de bénitier Tridacna noae, espèce dont le nom a été récemment ressuscité, pris en photo à Tiga (Iles Loyauté, Nouvelle-Calédonie) lors de la mission BIBELOT. On reconnaît T. noae aux dessins discrets en forme de larme (généralement entourés d'une bordure blanche) qui caractérisent son manteau (photo, Cécile Fauvelot).
Les échantillons de bénitiers récoltés lors de la mission BIBELOT ont été analysés et, ajoutés à d'autres échantillons récoltés dans l'Indo-Pacifique, ont permis de réaliser la première phylogénie complète des dix espèces connues de bénitiers du genre Tridacna basée sur une analyse multigénique calibrée à l'aide de fossiles, ceci afin de comprendre les relations phylogéniques entre espèces et retracer l'histoire évolutive de ce groupe (Fauvelot et al. 2020)
L'analyse génétique des échantillons de bénitiers récoltés est toujours en cours pour certaines espèces, mais nous avons déjà pu montrer, via les thèses des Josina Tiavouane et Simon van Wynsberge, que pour Tridacna maxima, Tridacna noae et Hippopus hippopus, les populations sont très structurées, avec une connectivité restreinte entre les populations des iles loyautés et celles du reste de la Nouvelle Calédonie (van Wynsberge et al. 2017, Fauvelot et al. 2019); Les résultats sur la connectivité des bénitiers de Nouvelle-Calédonie sont présentés dans un rapport d'Opération ZONECO, rédigé en français, disponible en cliquant sur le lien: DOI: 10.13140/RG.2.2.28827.21286
2) les Hydraires
Pour les hydraires collectés en Nouvelle-Calédonie et dans le monde, le marqueur mitochondrial 16S a été utilisé pour étudier la diversité des lignées chez les Aglaopheniidae et détecter les espèces cryptiques potentielles. Des analyses supplémentaires au sein de deux espèces sélectionnées, Lytocarpia brevirostris et Macrorhynchia phoenicea ont été réalisées à l'aide de marqueurs nucléaires microsatellites. Les méthodes de délimitation des espèces moléculaires et les analyses de génétique des populations ont révélé une grande diversité de lignées et plusieurs espèces cryptiques dans le groupe des Aglaopheniidae (Postaire et al. 2016, 2017a, 2017b). Ceci est illustré en détail, par exemple, chez l'espèce Lytocarpia brevirostris, avec des lignées alpha et ß trouvées en sympatrie sur les récifs de Nouvelle-Calédonie (Figure 2).
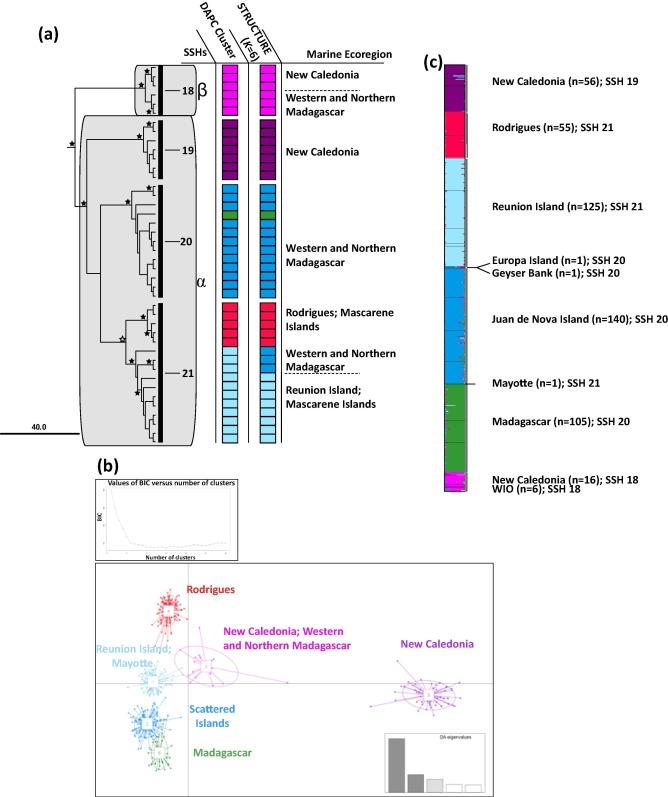
Figure 2: (a) Sous-arbre phylogénétique de Lytocarpia brevirostris réalisés à partir de trois gènes mitochondriaux et nucléaires avec les lignées alpha et ß et les hypothèses d'espèces secondaires (SSH) résultant des différentes méthodes de délimitation des espèces (barres verticales noires) avec (b) DAPC et (c) sorties STRUCTURE. (Figure de Postaire et al. (2016) https://doi.org/10.1016/j.ympev.2016.08.013)
Lorsque les échantillons ont été triés en fonction de leur appartenance à des lignées génétiquement différenciées, une analyse génétique supplémentaire au sein des lignées a montré une forte structuration parmi les populations échantillonnées (Figure 3), indiquant une dispersion larvaire limitée chez ces espèces (Postaire et al.2017a, 2017b).
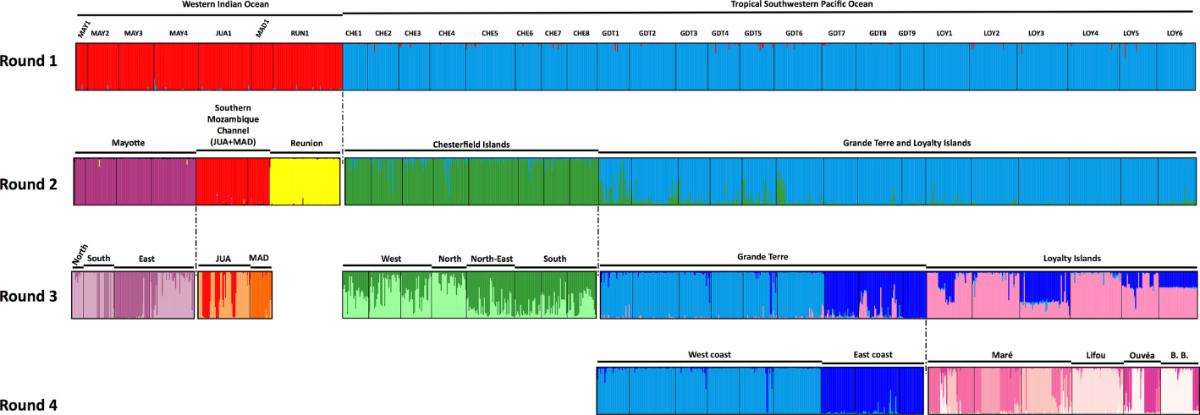
Figure 3. Structure génétique de la population dans la lignée alpha de Macrorhynchia phoenicea. Probabilités d'attribution des individus aux clusters supposés à partir de leurs génotypes multilocus aux marqueurs nucléaires microsatellites. Différentes couleurs correspondent à des clusters génétiquement différenciés. Figure de Postaire et al. (2017) https://doi.org/10.1002/ece3.3236
3) Coraux du genre Pocillopora
Chez les Pocilloporidae, des résultats similaires à ceux obtenus chez les hydraires ont été obtenus via les thèses de Pauline Gélin et Nicolas Oury (bourses doctorales financées par le LabEx CORAIL), à savoir un certain nombre de lignées cryptiques génétiquement différenciées, dont certaines présentes aux iles Loyautés (Gélin et al. 2017, 2018a, 2018b, Oury et al. 2020). Pour certaines des hypothèses d'espèces (SSH) trouvées dans le genre Pocillopora, d'autres lignées génétiquement distinctes ont été trouvées. C'est le cas dans le complexe d'espèces Pocillopora eydouxi / meandrina sensu, Schmidt-Roach, Miller, Lundgren et Andreakis (2014), identifié par Gélin, Postaire, Fauvelot et Magalon (2017) et qui s'est avéré se diviser en trois hypothèses d'espèces secondaires (SSH09a, SSH09b et SSH09c). Les SSH09b et SSH09c ont été trouvés presque exclusivement dans le Pacifique Sud tropical et le Pacifique Sud-Est (Figure 4). Plus surprenant, chaque SSH se divise en deux à trois clusters génétiquement différenciés, trouvés en sympatrie à l'échelle du récif (notamment en Nouvelle-Calédonie, y compris dans les îles Loyauté), conduisant à un modèle de niveaux hiérarchiques imbriqués (PSH> SSH> cluster), chacun niveau cachant des groupes génétiques très différenciés. Ainsi, plutôt que des populations structurées au sein d'une même espèce, ces trois SSH, et peut-être même les huit clusters, représentent probablement des lignées génétiques distinctes engagées dans un processus de spéciation (ou des espèces réelles). L'enjeu est maintenant de comprendre quel niveau hiérarchique (SSH, cluster, voire inférieur) correspond à celui de l'espèce. L'utilisation (en cours) de marqueurs moléculaires plus informatifs devrait nous aider à clarifier le statut taxonomique de ces lignées cryptiques.
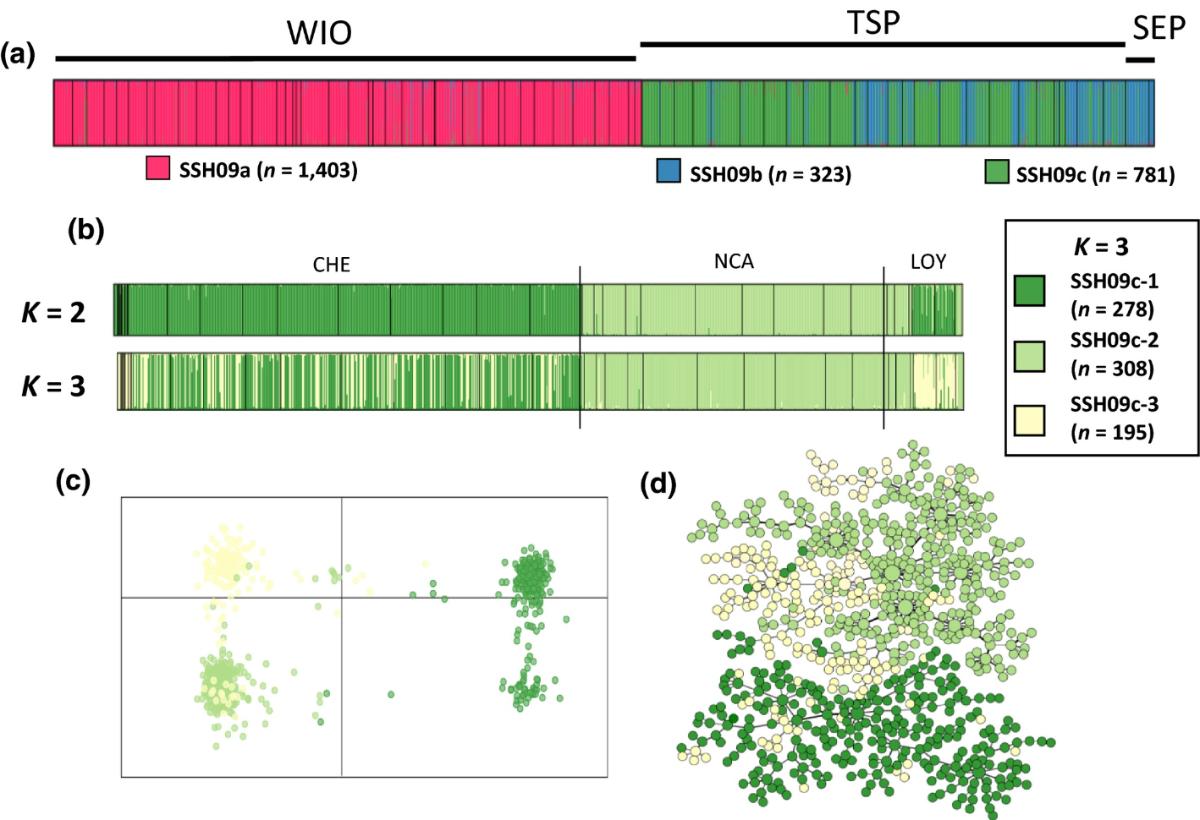
Figure 4: Variabilité génétique trouvée dans le cluster SSH09c du complexe d'espèces Pocillopora eydouxi / meandrina (cluster vert sur la figure 5a), à l'aide de marqueurs microsatellites. Chaque couleur correspond à des clusters génétiquement différenciés qui correspondent à des lignées génétiques divergentes que l'on retrouve en sympatrie sur les récifs de Chesterfield (CHE), de Nouvelle-Calédonie (NCA) et des îles Loyauté (LOY). (c) Résultats de l'affectation DAPC pour K = 3 et (d) réseau d'haplotypes, tous deux colorés selon les trois clusters identifiés par Structure. Article disponible à https://doi.org/10.1002/ece3.3747
L'ensemble de ces travaux représentent une avancée majeure dans le domaine puisque nous avons pu mettre en évidence un nombre insoupçonné d'espèces cryptiques chez ce genre, ce qui offre de nouvelles perspectives pour les autres genres de coraux où la taxonomie basée sur les caractères morphologiques est toujours incertaine et en constante révision. Des analyses complémentaires sont actuellement en cours dans le cadre de la thèse de Nicolas Oury.
4) Coraux d'espèces autres que ceux du genre Pocillopora
Les autres espèces de coraux ont majoritairement été collectées pour compléter la collection de coraux de Nouvelle Calédonie au centre IRD de Nouméa, mais on note quelques nouvelles identifications d'espèces récemment décrites chez les fungidae, avec notamment 2 espèces symbiotiques décrites pour la première fois : un serpulidae Spirobranchus richardsmithi (Pillai, 2009) sur un fungidae Podabacia motuporensis (Veron, 1990) qui a fait l'objet d'un article publié (Hoeksema & ten Hove 2014) et une crevette Ancylomenes kobayashii (Okuno & Nomura, 2002) sur un fungidae Polyphyllia novaehiberniae (Lesson, 1831). Aussi, de nombreuses espèces de coraux collectées aux iles Loyautés ont permis de compléter des phylogénies sur des groupes jusqu'alors méconnus (Arrigoni et al. 2016a, 2016b, 2019 ; Terraneo et al. 2014 ; van der Meij et al. 2015, Waheed et al. 2015).



